外側膝状体から1次視覚野へ入力する軸索の方位選択性は層により異なる
根東 覚・大木研一
(九州大学大学院医学研究院 分子生理学分野)
email:根東 覚,大木研一
DOI: 10.7875/first.author.2016.009
Laminar differences in the orientation selectivity of geniculate afferents in mouse primary visual cortex.
Satoru Kondo, Kenichi Ohki
Nature Neuroscience, 19, 316-319 (2016)
網膜において受容された外界の情報は視神経により外側膝状体へと伝達され,さらに大脳皮質の1次視覚野へと伝達される.1次視覚野のニューロンはさまざまな視覚の情報に対し選択的に反応しているが,そのなかのひとつに方位選択性がある.方位選択性とは特定の傾きの線にしか反応しない性質で,物体の輪郭など形の認識において重要な役割をはたすと考えられる.ネコやサルなど高等な哺乳類においては方位選択性をもつニューロンは網膜や外側膝状体にはほとんど存在せず,1次視覚野のニューロンの方位選択性は1次視覚野の神経回路において形成されると考えられている.ところが,マウスにおいては外側膝状体の一部のニューロンに方位選択性が報告されており,これが1次視覚野の方位選択性に対しどのような影響をおよぼしているのか問題になっていた.そこで,筆者らは,外側膝状体から1次視覚野への入力に方位選択性があるかどうか,外側膝状体から入力する軸索を1次視覚野において直接的に機能イメージングすることにより調べた.
マウスとネコやサルなど高等な哺乳類とでは1次視覚野の機能の構築に違いがある.方位選択性についてみると,高等な哺乳類には方位選択性の似たニューロンが層を貫いて集合する方位選択性コラムが認められるが,マウスなどげっ歯類にはそのような構造はない1).このような違いがあるにもかかわらず,1次視覚野のニューロンはいずれの生物種においても鋭い方位選択性をもつ.そこで,マウスと高等な哺乳類とのあいだで1次視覚野のニューロンが方位選択性を獲得する機構は保存されているのか,それとも,別の機構によるのかという疑問が生じる.方位選択性を形成する神経回路については,半世紀以上もまえに,1次視覚野のニューロンに方位選択性をもたない外側膝状体の円形の受容野がならぶように入力するという古典的なモデルが提唱され2),支持されてきている.しかしながら,マウスにおいて外側膝状体の一部のニューロンは方位選択性をもつことが報告されたことから3,4),マウスの1次視覚野のニューロンの方位選択性は高等な哺乳類とは異なる機構により形成される可能性がでてきた.
1次視覚野は大脳皮質の後頭葉にあり,深さにより6つの層に分類され,層ごとに機能が分化している.外側膝状体のニューロンからの投射をうける主要な層は第4層である.マウスの1次視覚野の第4層のニューロンに方位選択性が生じる機構については,外側膝状体から第4層への入力に方位選択性がありその入力がそのままうけわたされる,外側膝状体から第4層への入力それ自体の方位選択性は弱いがこの弱い方位選択性が発火の閾値により増幅されて方位選択性が生じる5),外側膝状体から第4層への入力には方位選択性がなく高等な哺乳類と同様に受容野の配列により方位選択性が生じる6),など,いくつかのモデルが提唱されている.これらのモデルは外側膝状体あるいは1次視覚野のニューロンからの活動の記録をもとに提唱されている.しかしながら,マウスの1次視覚野のニューロンが方位選択性を獲得する機構を理解するためには外側膝状体から1次視覚野の第4層へと入力する情報に方位選択性があるかどうかを知ることが不可欠と思われるが,この点については未解明であった.
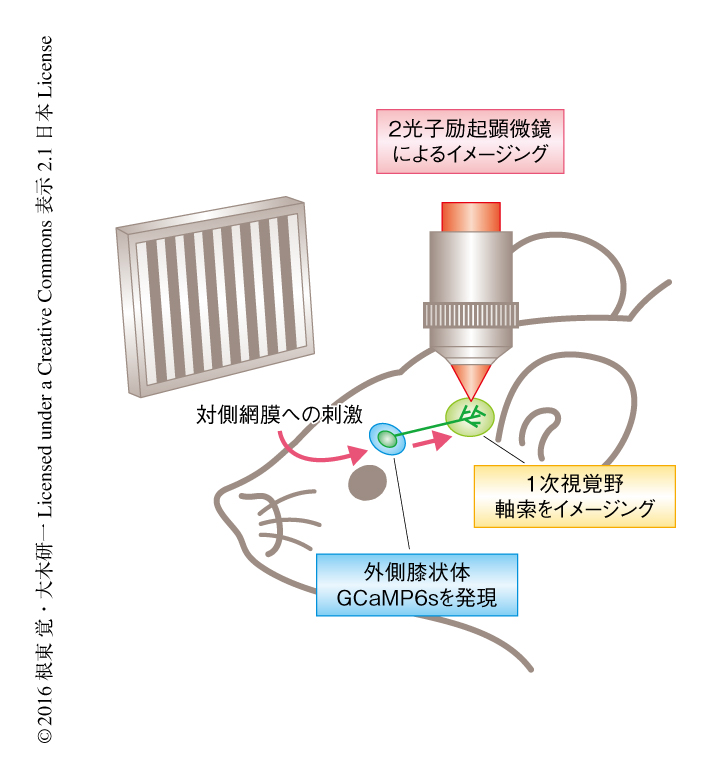
マウスの外側膝状体のニューロンが1次視覚野へ投射する軸索の活動を記録するため,神経活動の変化により蛍光の明るさが変化するCa2+感受性タンパク質GCaMP6sをアデノ随伴ウイルスベクターにより外側膝状体のニューロンのみに発現させた.アデノ随伴ウイルスベクターは非病原性であり,動物への遺伝子導入に適したウイルスベクターである.外側膝状体から1次視覚野へ投射する軸索の方位選択性は,2光子励起顕微鏡を用いてCa2+をイメージングすることにより調べた(図1).
2光子励起顕微鏡を用いて1次視覚野の第4層を観察したとき,脳の組織により発生する球面収差のため空間分解能が低下する.球面収差とは,対物レンズにより集めた光が光軸上において1点に集まらない現象である.脳の組織には細胞や細胞外組織など屈折率の異なる要素が多く混ざりあっているため,光の屈折する角度が一様にならず,光の収束する位置がずれることにより球面収差が発生し空間解像度が低下する.軸索は1μm程度の非常に細い小さな構造であるため,空間分解能の低下によりイメージングが困難になる.この球面収差を補正することにより,第1層と変わらない分解能で第4層に投射する軸索をイメージングすることが可能になった.その結果,外側膝状体から第4層に入力する軸索は方位選択性をあまりもたないことがわかった.一方,第4層よりも皮質の表面に近い第1層においては,第4層よりも高い割合で外側膝状体からの入力に方位選択性がみられた.また,すべての層において,方位選択性のみられた軸索について垂直あるいは水平な線に反応するものが多くみられた.
1次視覚野のニューロンは刺激を動かしたときの運動の方向に対する選択性をもつこともあり,これは方向選択性とよばれている.方位選択性をもつ1次視覚野のニューロンの一部は方向選択性ももつ.高等な哺乳類の外側膝状体のニューロンには方向選択性をもつものは非常に少ないが,げっ歯類およびウサギでは外側膝状体および網膜のニューロンにも方向選択性のあることが報告されている.最近,マウスの1次視覚野の第1層に方向選択性をもつ入力が報告され,この情報が網膜の方向選択性をもつニューロンから外側膝状体の背側部のシェルとよばれる特定の場所を経由して伝達されていることが示された7).そこで,方位選択性と同様の方法により外側膝状体の軸索の方向選択性を調べた結果,方位選択性と同様に,第4層に分布する軸索には方向選択性はほとんどみられなかったが,第1層には方向選択性をもつ軸索の入力が高い割合で認められた.
1次視覚野のニューロンは縞刺激の白と黒の間隔に対し選択的な反応を示す.この縞刺激の間隔を空間周波数とよび,ニューロンがもっともよく反応する間隔を最適空間周波数とよぶ.1次視覚野のニューロンは空間周波数に対する選択性をもつ.外側膝状体から入力する軸索の空間周波数に対する選択性を調べた結果,最適空間周波数の分布には層による大きな違いはみられなかった.
外側膝状体からの入力とそれをうけとる1次視覚野のニューロンとの反応選択性を比較したところ,いくつかの異なる点が明らかにされた.まず,方位選択性については,第4層に入力する外側膝状体の軸索の大部分には方位選択性がみられなかったのに対し,第4層のニューロンの大部分は鋭い方位選択性をもっていた.さらに,外側膝状体の軸索には方位選択性に垂直あるいは水平な線に顕著に反応するものがみられたが,1次視覚野のニューロンにはほとんどみられなかった.また,方向選択性についても方位選択性と同様の結果が得られた.空間周波数に対する選択性について最適空間周波数の分布をみると,外側膝状体の軸索は1次視覚野のニューロンより低い空間周波数に反応することがわかった.
外側膝状体から1次視覚野へ入力する軸索を1次視覚野のそれぞれの層において直接的にイメージングすることにより,それぞれの層に入力する情報を分けて調べることに成功した.その結果,第4層には方位選択性をもつ軸索はほとんど入力していないことがわかった.また,第1層などの皮質の表面に近い層には方位選択性をもつ入力が多くみられたことから,外側膝状体に存在する方位選択性をもつニューロンの多くは第4層以外に投射していることが示唆された.また,外側膝状体からの入力には方位選択性において垂直あるいは水平な線に顕著に反応するものがみられたが,第4層のニューロンにはほとんどみられなかった.空間周波数に対する選択性については,最適空間周波数の分布が外側膝状体からの入力と第4層のニューロンとのあいだで異なっていた.これらの結果から,外側膝状体の入力がそのまま1次視覚野のニューロンに伝達されるのではなく,大脳皮質の神経回路においてなんらかの計算が行われていることが示唆された.
層により反応選択性が異なるという結果は,外側膝状体のニューロンの単一の軸索のトレーシング解析からも支持されている.すなわち,おもに表層に軸索を送るタイプとおもに中間層に軸索を送るタイプに大別される.高等な哺乳類においても似たパターンが,大細胞経路,小細胞経路,顆粒細胞経路において報告されている8).表層に送られる方位選択的な情報の役割については,今後,明らかにする必要がある.
マウスも基本的にヒトと同じ神経回路により物を見ているとすると,マウスを用いてヒトが視覚の情報をどのような神経回路により処理しているのか,基本的な機構を理解することができるかもしれない.今回の結果は,視覚の情報が大脳において処理されるしくみの解明だけでなく,大脳の神経回路におけるより広く哺乳類に共通した情報処理の機構の理解にもつながることが期待される.
略歴:九州大学大学院医学研究院 助教.
大木 研一(Kenichi Ohki)
九州大学大学院医学研究院 教授.
研究室URL:http://www.physiol2.med.kyushu-u.ac.jp/
© 2016 根東 覚・大木研一 Licensed under CC 表示 2.1 日本
(九州大学大学院医学研究院 分子生理学分野)
email:根東 覚,大木研一
DOI: 10.7875/first.author.2016.009
Laminar differences in the orientation selectivity of geniculate afferents in mouse primary visual cortex.
Satoru Kondo, Kenichi Ohki
Nature Neuroscience, 19, 316-319 (2016)
要 約
網膜において受容された外界の情報は視神経により外側膝状体へと伝達され,さらに大脳皮質の1次視覚野へと伝達される.1次視覚野のニューロンはさまざまな視覚の情報に対し選択的に反応しているが,そのなかのひとつに方位選択性がある.方位選択性とは特定の傾きの線にしか反応しない性質で,物体の輪郭など形の認識において重要な役割をはたすと考えられる.ネコやサルなど高等な哺乳類においては方位選択性をもつニューロンは網膜や外側膝状体にはほとんど存在せず,1次視覚野のニューロンの方位選択性は1次視覚野の神経回路において形成されると考えられている.ところが,マウスにおいては外側膝状体の一部のニューロンに方位選択性が報告されており,これが1次視覚野の方位選択性に対しどのような影響をおよぼしているのか問題になっていた.そこで,筆者らは,外側膝状体から1次視覚野への入力に方位選択性があるかどうか,外側膝状体から入力する軸索を1次視覚野において直接的に機能イメージングすることにより調べた.
はじめに
マウスとネコやサルなど高等な哺乳類とでは1次視覚野の機能の構築に違いがある.方位選択性についてみると,高等な哺乳類には方位選択性の似たニューロンが層を貫いて集合する方位選択性コラムが認められるが,マウスなどげっ歯類にはそのような構造はない1).このような違いがあるにもかかわらず,1次視覚野のニューロンはいずれの生物種においても鋭い方位選択性をもつ.そこで,マウスと高等な哺乳類とのあいだで1次視覚野のニューロンが方位選択性を獲得する機構は保存されているのか,それとも,別の機構によるのかという疑問が生じる.方位選択性を形成する神経回路については,半世紀以上もまえに,1次視覚野のニューロンに方位選択性をもたない外側膝状体の円形の受容野がならぶように入力するという古典的なモデルが提唱され2),支持されてきている.しかしながら,マウスにおいて外側膝状体の一部のニューロンは方位選択性をもつことが報告されたことから3,4),マウスの1次視覚野のニューロンの方位選択性は高等な哺乳類とは異なる機構により形成される可能性がでてきた.
1次視覚野は大脳皮質の後頭葉にあり,深さにより6つの層に分類され,層ごとに機能が分化している.外側膝状体のニューロンからの投射をうける主要な層は第4層である.マウスの1次視覚野の第4層のニューロンに方位選択性が生じる機構については,外側膝状体から第4層への入力に方位選択性がありその入力がそのままうけわたされる,外側膝状体から第4層への入力それ自体の方位選択性は弱いがこの弱い方位選択性が発火の閾値により増幅されて方位選択性が生じる5),外側膝状体から第4層への入力には方位選択性がなく高等な哺乳類と同様に受容野の配列により方位選択性が生じる6),など,いくつかのモデルが提唱されている.これらのモデルは外側膝状体あるいは1次視覚野のニューロンからの活動の記録をもとに提唱されている.しかしながら,マウスの1次視覚野のニューロンが方位選択性を獲得する機構を理解するためには外側膝状体から1次視覚野の第4層へと入力する情報に方位選択性があるかどうかを知ることが不可欠と思われるが,この点については未解明であった.
1.外側膝状体から1次視覚野へ入力する軸索の方位選択性
マウスの外側膝状体のニューロンが1次視覚野へ投射する軸索の活動を記録するため,神経活動の変化により蛍光の明るさが変化するCa2+感受性タンパク質GCaMP6sをアデノ随伴ウイルスベクターにより外側膝状体のニューロンのみに発現させた.アデノ随伴ウイルスベクターは非病原性であり,動物への遺伝子導入に適したウイルスベクターである.外側膝状体から1次視覚野へ投射する軸索の方位選択性は,2光子励起顕微鏡を用いてCa2+をイメージングすることにより調べた(図1).
2光子励起顕微鏡を用いて1次視覚野の第4層を観察したとき,脳の組織により発生する球面収差のため空間分解能が低下する.球面収差とは,対物レンズにより集めた光が光軸上において1点に集まらない現象である.脳の組織には細胞や細胞外組織など屈折率の異なる要素が多く混ざりあっているため,光の屈折する角度が一様にならず,光の収束する位置がずれることにより球面収差が発生し空間解像度が低下する.軸索は1μm程度の非常に細い小さな構造であるため,空間分解能の低下によりイメージングが困難になる.この球面収差を補正することにより,第1層と変わらない分解能で第4層に投射する軸索をイメージングすることが可能になった.その結果,外側膝状体から第4層に入力する軸索は方位選択性をあまりもたないことがわかった.一方,第4層よりも皮質の表面に近い第1層においては,第4層よりも高い割合で外側膝状体からの入力に方位選択性がみられた.また,すべての層において,方位選択性のみられた軸索について垂直あるいは水平な線に反応するものが多くみられた.
2.外側膝状体から1次視覚野へと入力する軸索の方向選択性
1次視覚野のニューロンは刺激を動かしたときの運動の方向に対する選択性をもつこともあり,これは方向選択性とよばれている.方位選択性をもつ1次視覚野のニューロンの一部は方向選択性ももつ.高等な哺乳類の外側膝状体のニューロンには方向選択性をもつものは非常に少ないが,げっ歯類およびウサギでは外側膝状体および網膜のニューロンにも方向選択性のあることが報告されている.最近,マウスの1次視覚野の第1層に方向選択性をもつ入力が報告され,この情報が網膜の方向選択性をもつニューロンから外側膝状体の背側部のシェルとよばれる特定の場所を経由して伝達されていることが示された7).そこで,方位選択性と同様の方法により外側膝状体の軸索の方向選択性を調べた結果,方位選択性と同様に,第4層に分布する軸索には方向選択性はほとんどみられなかったが,第1層には方向選択性をもつ軸索の入力が高い割合で認められた.
3.外側膝状体から1次視覚野へと入力する軸索の空間周波数に対する選択性
1次視覚野のニューロンは縞刺激の白と黒の間隔に対し選択的な反応を示す.この縞刺激の間隔を空間周波数とよび,ニューロンがもっともよく反応する間隔を最適空間周波数とよぶ.1次視覚野のニューロンは空間周波数に対する選択性をもつ.外側膝状体から入力する軸索の空間周波数に対する選択性を調べた結果,最適空間周波数の分布には層による大きな違いはみられなかった.
4.外側膝状体の軸索と1次視覚野のニューロンとの反応選択性の比較
外側膝状体からの入力とそれをうけとる1次視覚野のニューロンとの反応選択性を比較したところ,いくつかの異なる点が明らかにされた.まず,方位選択性については,第4層に入力する外側膝状体の軸索の大部分には方位選択性がみられなかったのに対し,第4層のニューロンの大部分は鋭い方位選択性をもっていた.さらに,外側膝状体の軸索には方位選択性に垂直あるいは水平な線に顕著に反応するものがみられたが,1次視覚野のニューロンにはほとんどみられなかった.また,方向選択性についても方位選択性と同様の結果が得られた.空間周波数に対する選択性について最適空間周波数の分布をみると,外側膝状体の軸索は1次視覚野のニューロンより低い空間周波数に反応することがわかった.
おわりに
外側膝状体から1次視覚野へ入力する軸索を1次視覚野のそれぞれの層において直接的にイメージングすることにより,それぞれの層に入力する情報を分けて調べることに成功した.その結果,第4層には方位選択性をもつ軸索はほとんど入力していないことがわかった.また,第1層などの皮質の表面に近い層には方位選択性をもつ入力が多くみられたことから,外側膝状体に存在する方位選択性をもつニューロンの多くは第4層以外に投射していることが示唆された.また,外側膝状体からの入力には方位選択性において垂直あるいは水平な線に顕著に反応するものがみられたが,第4層のニューロンにはほとんどみられなかった.空間周波数に対する選択性については,最適空間周波数の分布が外側膝状体からの入力と第4層のニューロンとのあいだで異なっていた.これらの結果から,外側膝状体の入力がそのまま1次視覚野のニューロンに伝達されるのではなく,大脳皮質の神経回路においてなんらかの計算が行われていることが示唆された.
層により反応選択性が異なるという結果は,外側膝状体のニューロンの単一の軸索のトレーシング解析からも支持されている.すなわち,おもに表層に軸索を送るタイプとおもに中間層に軸索を送るタイプに大別される.高等な哺乳類においても似たパターンが,大細胞経路,小細胞経路,顆粒細胞経路において報告されている8).表層に送られる方位選択的な情報の役割については,今後,明らかにする必要がある.
マウスも基本的にヒトと同じ神経回路により物を見ているとすると,マウスを用いてヒトが視覚の情報をどのような神経回路により処理しているのか,基本的な機構を理解することができるかもしれない.今回の結果は,視覚の情報が大脳において処理されるしくみの解明だけでなく,大脳の神経回路におけるより広く哺乳類に共通した情報処理の機構の理解にもつながることが期待される.
文 献
- Ohki, K., Chung, S., Ch’ng, Y. H. et al.: Functional imaging with cellular resolution reveals precise micro-architecture in visual cortex. Nature, 433, 597-603 (2005)[PubMed]
- Hubel, D. H. & Wiesel, T. N.: Receptive fields, binocular interaction and functional architecture in the cat’s visual cortex. J. Physiol., 160, 106-154 (1962)[PubMed]
- Marshel, J. H., Kaye, A. P., Nauhaus, I. et al.: Anterior-posterior direction opponency in the superficial mouse lateral geniculate nucleus. Neuron, 76, 713-720 (2012)[PubMed]
- Piscopo, D. M., El-Danaf, R. N., Huberman, A. D. et al.: Diverse visual features encoded in mouse lateral geniculate nucleus. J. Neurosci., 33, 4642-4656 (2013)[PubMed]
- Scholl, B., Tan, A. Y., Corey, J. et al.: Emergence of orientation selectivity in the mammalian visual pathway. J. Neurosci., 33, 10616-10624 (2013)[PubMed]
- Lien, A. D. & Scanziani, M.: Tuned thalamic excitation is amplified by visual cortical circuits. Nat. Neurosci., 16, 1315-1323 (2013)[PubMed]
- Cruz-Martin, A., El-Danaf, R. N., Osakada, F. et al.: A dedicated circuit links direction-selective retinal ganglion cells to the primary visual cortex. Nature, 507, 358-361 (2014)[PubMed]
- Lund, J. S.: Anatomical organization of macaque monkey striate visual cortex. Annu. Rev. Neurosci., 253-288 (1988)[PubMed]
著者プロフィール
略歴:九州大学大学院医学研究院 助教.
大木 研一(Kenichi Ohki)
九州大学大学院医学研究院 教授.
研究室URL:http://www.physiol2.med.kyushu-u.ac.jp/
© 2016 根東 覚・大木研一 Licensed under CC 表示 2.1 日本
