ソマトスタチン陽性ニューロンは音の刺激の重要性に応じ1次聴覚野における応答を双方向性に制御する
加藤紘之・Jeffry S. Isaacson
(米国California大学San Diego校Department of Neurosciences)
email:加藤紘之
DOI: 10.7875/first.author.2015.130
Flexible sensory representations in auditory cortex driven by behavioral relevance.
Hiroyuki K. Kato, Shea N. Gillet, Jeffry S. Isaacson
Neuron, 88, 1027-1039 (2015)
生物における感覚の情報処理は固定されたものではなく,同一の入力であっても意味をともなわない場合にはそれを無視し,生存にかかわる場合には即座に反応することが可能になっている.この研究においては,このような状況に応じた情報処理の神経回路レベルでの基盤を明らかにするため,受動的な音の経験のくり返しによる馴化,および,音を用いたタスクへの従事の2つの過程において,大脳の1次聴覚野の応答性がどのように変化するか検討した.細胞種に特異的なCa2+イメージング法の結果,受動的な音の経験は1次聴覚野におけるソマトスタチン陽性ニューロンの神経活動を上昇させ,結果として,2/3層の錐体細胞において音への応答性を低下させた.これに対し,音を用いたタスクに従事しているあいだはソマトスタチン陽性ニューロンの神経活動は低下し,2/3層の錐体細胞において音への応答性を上昇させた.これらの実験により,受動的な音の経験のくり返しによる知覚の低下,および,能動的な注意による知覚の向上という双方向性の知覚の制御における神経回路レベルでの基盤が明らかにされた.ソマトスタチン陽性ニューロンは感覚の刺激の重要性に応じ脳における情報の流れを制御するゲートとしての役割をもつと考えられた.
感覚の情報をその重要性に応じ適切に処理して行動を出力することは生物の生存において必須である.個々の感覚入力のもつ重要性は生来的には決定されておらず,それぞれの生物が得た経験に応じ更新されつづける動的なものである.このような重要性の変動に応じ行動の出力を変化させる役割の一端を担うと考えられているのが,大脳の感覚野における神経活動の可塑性である.実際に,大脳の1次聴覚野においては生育の初期に多く経験した周波数の音に応答する脳の領域が拡大することが知られている1,2).それに対し,成熟した動物の1次聴覚野はこのような単純な経験による変化にとぼしく,可塑性の誘導には電気ショックや報酬などとの関連づけが必要であると考えられてきた3,4).しかし,これら生理学的な知見とは対照的に,ヒトにおける心理学的な実験からは,受動的な音の刺激のくり返しは知覚される音量の低下(馴化)をひき起こすことが知られている5).このように,麻酔した動物における生理学のレベルでの知見とヒトにおける知覚のレベルでの知見には不一致があり,知覚のレベルにおける馴化の基盤となる可塑性の存在については明らかにされていなかった.
この研究においては,受動的な音の経験が1次聴覚野にあたえる影響を覚醒したマウスにおいて直接的に評価することを目的として,生体内Ca2+イメージング法により神経活動を長期にわたり記録した.さらに,マウスが音を用いたタスクに従事しているあいだの神経活動をあわせて観察することにより,音の重要性の低下(受動的な経験)および上昇(タスクへの従事)の2種類の過程での1次聴覚野における応答性の変化,および,その神経回路レベルでの機構の解明をめざした.
これまで,1次聴覚野における音への応答の測定には,ほとんどの場合,麻酔した動物に対する電気生理学的な手法が用いられてきた.しかしながら,神経回路における情報処理は覚醒時と麻酔時とでは大きく異なることが知られている6,7).そこで,覚醒した動物において音への応答を測定するため,Ca2+のインジケーターであるGCaMP6sをウイルスベクターを用いて1次聴覚野に発現させた.GCaMP6sを発現した細胞において,神経活動の変化はその蛍光強度の増加および減少として観察される.さらに,頭骨に観察用のガラス窓を埋めこみ2光子励起顕微鏡のもと頭部を固定することにより,覚醒したマウスにおいて同一の細胞の集団を長期にわたり安定して観察することが可能になった.大脳皮質には興奮性ニューロンである錐体細胞だけでなく,固有の役割をもつ多様な抑制性ニューロンの存在が知られている8).そこで,すべての抑制性ニューロンにおいて蛍光タンパク質であるtdTomatoを発現するトランスジェニックマウスを用いることにより,神経活動を錐体細胞から選択的に測定した.これらのマウスに5~9秒間にわたり継続する純音を聞かせたところ,1次聴覚野の2/3層の錐体細胞は興奮性および抑制性を含む多様な応答を示した.そこで,この応答が受動的な音の経験によりどのように変化するかについて検討するため,さらに同じ音の刺激を1日に200回,5日間にわたり聞かせたところ,錐体細胞における興奮性の応答は日をおって減少した(図1a).この興奮性の低下は知覚のレベルでの馴化に対応するものであり,これまでの麻酔した動物における報告とは異なり,覚醒した動物においては神経活動にも馴化が誘導されることが明らかにされた.興奮性の応答の減少と並行して,抑制性の応答を示す錐体細胞の割合は上昇した.この結果から,1次聴覚野における馴化は単純な応答性の低下ではなく,抑制性の入力の積極的な増加によることが示唆された.
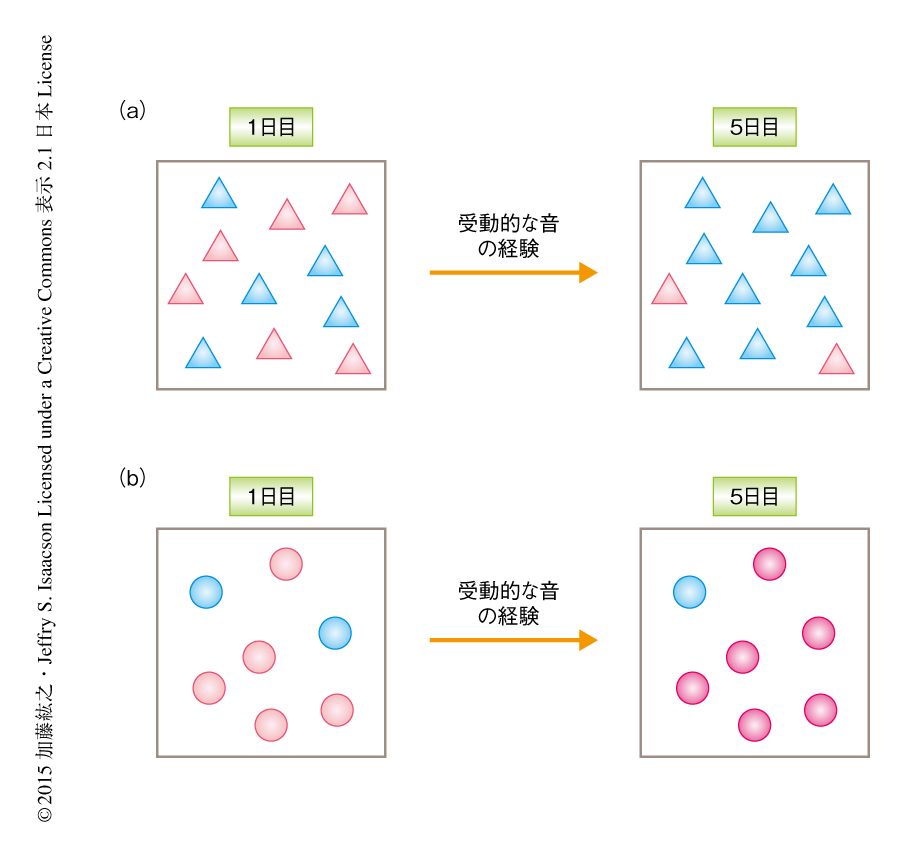
1次聴覚野の2/3層の錐体細胞において観察された音への応答の馴化はどのような神経回路により誘導されるのだろうか? この課題を検討するため,さまざまな細胞種にてリコンビナーゼであるCreを発現するトランスジェニックマウスを利用し,Ca2+インジケーターGCaMP6sをCreに依存して発現させることによりCa2+イメージング法を細胞種に選択的に行った.感覚の刺激の入力層である1次聴覚野の4層における興奮性ニューロンの神経活動を測定したところ,2/3層とは異なり,5日間にわたる受動的な音の経験は興奮性の応答のわずかな減少をひき起こしたのみで,抑制性の応答の増加は観察されなかった.この結果から,1次聴覚野における馴化はより上流に位置する視床からひきつがれたものではなく,2/3層それ自体において誘導されることが示された.そこで,錐体細胞への抑制性の入力の大部分を担うパルブアルブミン陽性ニューロンおよびソマトスタチン陽性ニューロンに着目し同様に実験した.その結果,錐体細胞の場合とは逆に,ソマトスタチン陽性ニューロンの興奮性の応答は受動的な音の経験により日をおって増加した(図1b).もうひとつの主要な抑制性ニューロンであるパルブアルブミン陽性ニューロンにおいては,錐体細胞と同様の興奮性の応答の減少が観察された.したがって,受動的な音の刺激のくり返しにより誘導される錐体細胞への抑制性の入力の増加は,ソマトスタチン陽性ニューロンに依存することが示唆された.これらの結果から,知覚における馴化の神経回路レベルでの基盤が明らかにされるとともに,特定の抑制性ニューロンがこの可塑性に寄与している可能性が示された.
以上の結果から,意味をともなわない受動的な音の刺激のくり返しが1次聴覚野にひき起こす可塑性が明らかにされた.それでは逆に,音の刺激に意味があたえられた場合にはどのような変化が起こるのだろうか? 受動的な経験が音に対する知覚を低下させるのとは対照的に,特定の音への能動的な注意はその知覚を向上させることが知られている9).しかしながら,注意にともなう大脳の1次聴覚野における神経活動の変化について,一定の見解は得られていない10-12).そこで,知覚の向上の神経回路レベルでの基盤について明らかにするため,頭部を固定したマウスを音の刺激を用いたタスクに従事させ,1次聴覚野における音への応答性の変化について検討した.一般的な行動実験では音の刺激の開始を報告させるタスクが多く用いられているが,これらの実験には1次聴覚野における音への応答が報告にともなう運動の影響をうけるという問題点があった13).そこでこの実験においては,マウスに5~9秒間にわたり継続する音の刺激をあたえ,刺激の開始ではなく終了のタイミングを報告した場合に報酬をあたえた.このタスクにおいては音の刺激のあいだマウスは静止しているため,運動による影響をうけることなく音への応答を測定することが可能になった.
まず,1次聴覚野の2/3層の錐体細胞における神経活動をCa2+イメージング法により測定し,同一の細胞において,マウスがタスクに従事しているあいだの音への応答(行動時)と,受動的に音を聞いているあいだの音への応答(受動時)とを比較したところ,行動時においては錐体細胞の興奮性の応答が受動時よりも増加していた.この結果は,能動的な注意により知覚が向上するという心理学的な知見と一致するものであった.そこで,この可塑性の神経回路レベルでの基盤について明らかにするため,ふたたびトランスジェニックマウスを用いてCa2+イメージング法を細胞種に選択的に行った.その結果,1次聴覚野の4層の興奮性ニューロンにおける神経活動は行動時と受動時とで変化はなく,この可塑性が2/3層それ自体において誘導されていることが示された.パルブアルブミン陽性ニューロンおよびソマトスタチン陽性ニューロンにおいて同様に実験したところ,錐体細胞の場合とは逆に,ソマトスタチン陽性ニューロンの興奮性の応答のみが行動時において減少していた.これらの結果から,行動時においてはソマトスタチン陽性ニューロンにおける神経活動の低下が2/3層における錐体細胞の脱抑制をひき起こし,聴覚野における音への応答性を上昇させることが示唆された.
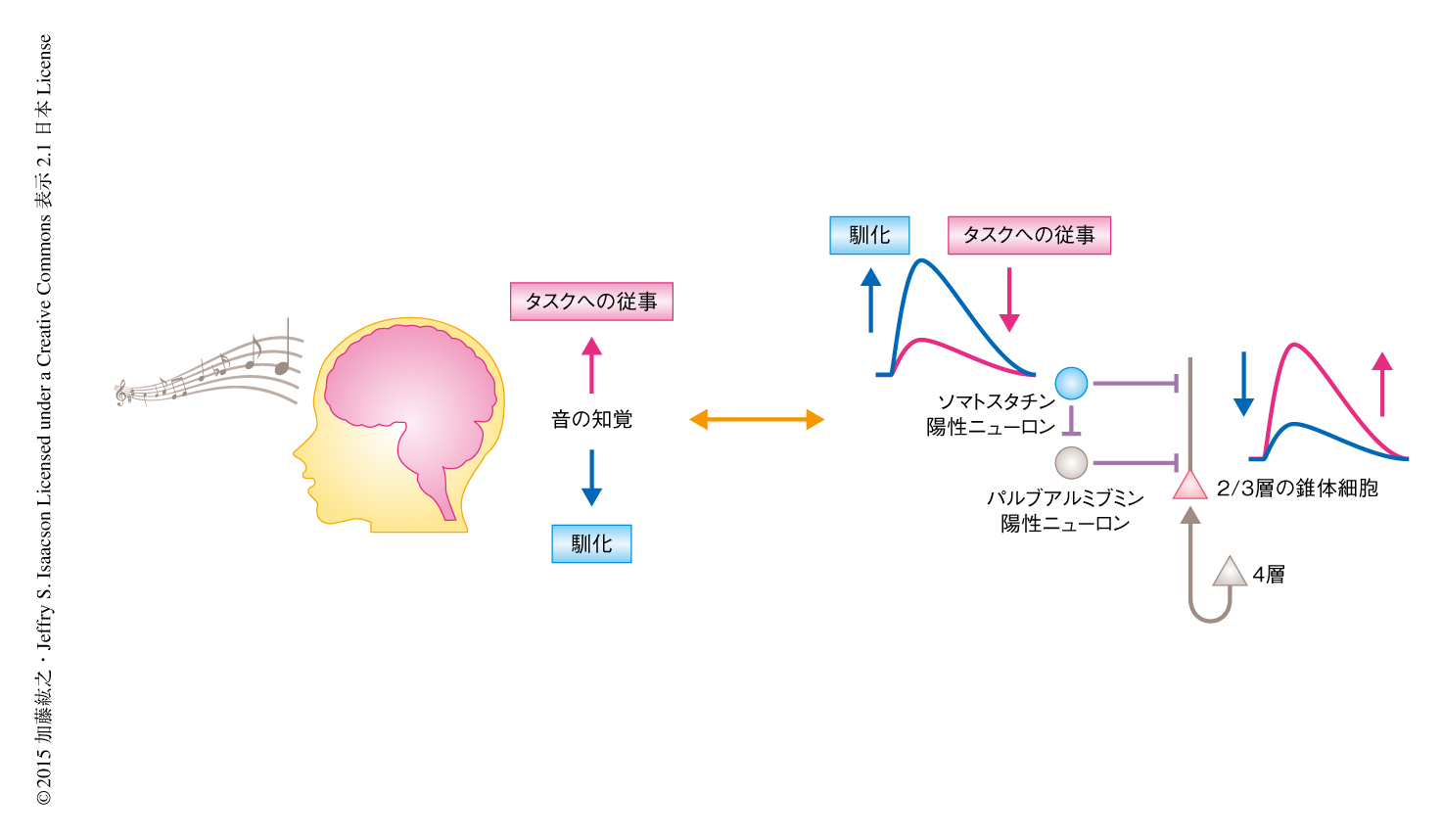
この研究においては,マウスにおける長期にわたる生体内Ca2+イメージング法を用いて,受動的な音の経験のくり返しが1次聴覚野の2/3層における錐体細胞の音への応答性を日をおって低下させることを見い出した.逆に,マウスが音の刺激を用いたタスクに従事しているあいだは錐体細胞の音への応答性の一時的な上昇が観察された.したがって,1次聴覚野における音への応答は成熟した動物においても固定されたものではなく,音の刺激のもつ重要性に応じ双方向性に制御されることが明らかにされた.さらに,これらの双方向性の制御はいずれもソマトスタチン陽性ニューロンによる抑制の変化をともなっていた.これらの結果から,ソマトスタチン陽性ニューロンは感覚の刺激の重要性に応じ1次聴覚野における情報の流れを制御するゲートとしての役割をもつと考えられた(図2).
これまでの多くの研究において,成熟した動物の1次聴覚野における音への応答性は固定されたものであり,受動的な音の経験は可塑性を誘導しないと報告されてきた1,3,4).これらの報告とは異なり,この研究において可塑性が観察された理由として2つの可能性が考えられた.第1に,細胞外記録を用いた以前の電気生理学的な実験においては神経活動がどの細胞種に由来するのか不明であった点があげられる.この研究においては,2/3層の錐体細胞,4層の興奮性ニューロン,ソマトスタチン陽性ニューロンがそれぞれ異なる可塑性を示すことが明らかにされた.細胞外記録は高い神経活動を示す細胞からの記録にかたよる傾向が強いため,以前の研究においては,受動的な音の経験ののちにも馴化を起こさない4層の細胞やソマトスタチン陽性ニューロンから記録をとっていた可能性がある.第2に,この研究が覚醒したマウスを用いたのとは対照的に,以前の多くの実験は麻酔のもとであった点があげられる.脳は覚醒時と麻酔時において異なる神経活動のパターンを示すが,とくに,抑制性ニューロンの神経活動は麻酔のもとでは大きく低下することが脳の複数の部位において報告されている6,7).したがって,麻酔のもとでは抑制性のソマトスタチン陽性ニューロンの神経活動が低下し,結果として,可塑性の発現が抑えられた可能性が高い.
この研究では,1次聴覚野における局所的な神経回路に着目し,その可塑性におけるソマトスタチン陽性ニューロンの重要性を示した.それでは,ソマトスタチン陽性ニューロンそれ自体の神経活動はどのような機構により制御されているのだろうか.その可能性として,ノルアドレナリンやアセチルコリンなどの神経修飾物質,あるいは,前頭皮質など高次の領域からの入力といった,脳の領域をまたいだ相互作用の寄与が考えられる.これらの経路を光遺伝学などにより操作し,ソマトスタチン陽性ニューロンの神経活動,さらに,1次聴覚野の可塑性にどのような影響がみられるかを検討することが今後の課題である.将来的には,これまでの局所的な神経回路の研究から1歩進めて,局所の神経回路どうしをつなぐより大きなネットワークを遺伝学的なツールを用いて切り分けることにより,脳がいかにして状況に応じた情報処理をするのか,より包括的な理解を進めたい.
略歴:2008年 東京大学大学院医学系研究科博士課程 修了,同年 同 博士研究員を経て,2010年より米国California大学San Diego校 博士研究員.
研究テーマ:生体における2光子励起イメージング法を用いた聴覚の情報処理の機構の解析.
Jeffry S. Isaacson
米国California大学San Diego校Professor.
研究室URL:http://isaacsonlab.net/Home.html
© 2015 加藤紘之・Jeffry S. Isaacson Licensed under CC 表示 2.1 日本
(米国California大学San Diego校Department of Neurosciences)
email:加藤紘之
DOI: 10.7875/first.author.2015.130
Flexible sensory representations in auditory cortex driven by behavioral relevance.
Hiroyuki K. Kato, Shea N. Gillet, Jeffry S. Isaacson
Neuron, 88, 1027-1039 (2015)
要 約
生物における感覚の情報処理は固定されたものではなく,同一の入力であっても意味をともなわない場合にはそれを無視し,生存にかかわる場合には即座に反応することが可能になっている.この研究においては,このような状況に応じた情報処理の神経回路レベルでの基盤を明らかにするため,受動的な音の経験のくり返しによる馴化,および,音を用いたタスクへの従事の2つの過程において,大脳の1次聴覚野の応答性がどのように変化するか検討した.細胞種に特異的なCa2+イメージング法の結果,受動的な音の経験は1次聴覚野におけるソマトスタチン陽性ニューロンの神経活動を上昇させ,結果として,2/3層の錐体細胞において音への応答性を低下させた.これに対し,音を用いたタスクに従事しているあいだはソマトスタチン陽性ニューロンの神経活動は低下し,2/3層の錐体細胞において音への応答性を上昇させた.これらの実験により,受動的な音の経験のくり返しによる知覚の低下,および,能動的な注意による知覚の向上という双方向性の知覚の制御における神経回路レベルでの基盤が明らかにされた.ソマトスタチン陽性ニューロンは感覚の刺激の重要性に応じ脳における情報の流れを制御するゲートとしての役割をもつと考えられた.
はじめに
感覚の情報をその重要性に応じ適切に処理して行動を出力することは生物の生存において必須である.個々の感覚入力のもつ重要性は生来的には決定されておらず,それぞれの生物が得た経験に応じ更新されつづける動的なものである.このような重要性の変動に応じ行動の出力を変化させる役割の一端を担うと考えられているのが,大脳の感覚野における神経活動の可塑性である.実際に,大脳の1次聴覚野においては生育の初期に多く経験した周波数の音に応答する脳の領域が拡大することが知られている1,2).それに対し,成熟した動物の1次聴覚野はこのような単純な経験による変化にとぼしく,可塑性の誘導には電気ショックや報酬などとの関連づけが必要であると考えられてきた3,4).しかし,これら生理学的な知見とは対照的に,ヒトにおける心理学的な実験からは,受動的な音の刺激のくり返しは知覚される音量の低下(馴化)をひき起こすことが知られている5).このように,麻酔した動物における生理学のレベルでの知見とヒトにおける知覚のレベルでの知見には不一致があり,知覚のレベルにおける馴化の基盤となる可塑性の存在については明らかにされていなかった.
この研究においては,受動的な音の経験が1次聴覚野にあたえる影響を覚醒したマウスにおいて直接的に評価することを目的として,生体内Ca2+イメージング法により神経活動を長期にわたり記録した.さらに,マウスが音を用いたタスクに従事しているあいだの神経活動をあわせて観察することにより,音の重要性の低下(受動的な経験)および上昇(タスクへの従事)の2種類の過程での1次聴覚野における応答性の変化,および,その神経回路レベルでの機構の解明をめざした.
1.受動的な音の経験のくり返しは1次聴覚野における応答性を長期的に低下させる
これまで,1次聴覚野における音への応答の測定には,ほとんどの場合,麻酔した動物に対する電気生理学的な手法が用いられてきた.しかしながら,神経回路における情報処理は覚醒時と麻酔時とでは大きく異なることが知られている6,7).そこで,覚醒した動物において音への応答を測定するため,Ca2+のインジケーターであるGCaMP6sをウイルスベクターを用いて1次聴覚野に発現させた.GCaMP6sを発現した細胞において,神経活動の変化はその蛍光強度の増加および減少として観察される.さらに,頭骨に観察用のガラス窓を埋めこみ2光子励起顕微鏡のもと頭部を固定することにより,覚醒したマウスにおいて同一の細胞の集団を長期にわたり安定して観察することが可能になった.大脳皮質には興奮性ニューロンである錐体細胞だけでなく,固有の役割をもつ多様な抑制性ニューロンの存在が知られている8).そこで,すべての抑制性ニューロンにおいて蛍光タンパク質であるtdTomatoを発現するトランスジェニックマウスを用いることにより,神経活動を錐体細胞から選択的に測定した.これらのマウスに5~9秒間にわたり継続する純音を聞かせたところ,1次聴覚野の2/3層の錐体細胞は興奮性および抑制性を含む多様な応答を示した.そこで,この応答が受動的な音の経験によりどのように変化するかについて検討するため,さらに同じ音の刺激を1日に200回,5日間にわたり聞かせたところ,錐体細胞における興奮性の応答は日をおって減少した(図1a).この興奮性の低下は知覚のレベルでの馴化に対応するものであり,これまでの麻酔した動物における報告とは異なり,覚醒した動物においては神経活動にも馴化が誘導されることが明らかにされた.興奮性の応答の減少と並行して,抑制性の応答を示す錐体細胞の割合は上昇した.この結果から,1次聴覚野における馴化は単純な応答性の低下ではなく,抑制性の入力の積極的な増加によることが示唆された.
2.1次聴覚野における馴化はソマトスタチン陽性ニューロンの神経活動の亢進をともなう
1次聴覚野の2/3層の錐体細胞において観察された音への応答の馴化はどのような神経回路により誘導されるのだろうか? この課題を検討するため,さまざまな細胞種にてリコンビナーゼであるCreを発現するトランスジェニックマウスを利用し,Ca2+インジケーターGCaMP6sをCreに依存して発現させることによりCa2+イメージング法を細胞種に選択的に行った.感覚の刺激の入力層である1次聴覚野の4層における興奮性ニューロンの神経活動を測定したところ,2/3層とは異なり,5日間にわたる受動的な音の経験は興奮性の応答のわずかな減少をひき起こしたのみで,抑制性の応答の増加は観察されなかった.この結果から,1次聴覚野における馴化はより上流に位置する視床からひきつがれたものではなく,2/3層それ自体において誘導されることが示された.そこで,錐体細胞への抑制性の入力の大部分を担うパルブアルブミン陽性ニューロンおよびソマトスタチン陽性ニューロンに着目し同様に実験した.その結果,錐体細胞の場合とは逆に,ソマトスタチン陽性ニューロンの興奮性の応答は受動的な音の経験により日をおって増加した(図1b).もうひとつの主要な抑制性ニューロンであるパルブアルブミン陽性ニューロンにおいては,錐体細胞と同様の興奮性の応答の減少が観察された.したがって,受動的な音の刺激のくり返しにより誘導される錐体細胞への抑制性の入力の増加は,ソマトスタチン陽性ニューロンに依存することが示唆された.これらの結果から,知覚における馴化の神経回路レベルでの基盤が明らかにされるとともに,特定の抑制性ニューロンがこの可塑性に寄与している可能性が示された.
3.音の刺激を用いたタスクへの従事は1次聴覚野における音への応答性を一時的に上昇させる
以上の結果から,意味をともなわない受動的な音の刺激のくり返しが1次聴覚野にひき起こす可塑性が明らかにされた.それでは逆に,音の刺激に意味があたえられた場合にはどのような変化が起こるのだろうか? 受動的な経験が音に対する知覚を低下させるのとは対照的に,特定の音への能動的な注意はその知覚を向上させることが知られている9).しかしながら,注意にともなう大脳の1次聴覚野における神経活動の変化について,一定の見解は得られていない10-12).そこで,知覚の向上の神経回路レベルでの基盤について明らかにするため,頭部を固定したマウスを音の刺激を用いたタスクに従事させ,1次聴覚野における音への応答性の変化について検討した.一般的な行動実験では音の刺激の開始を報告させるタスクが多く用いられているが,これらの実験には1次聴覚野における音への応答が報告にともなう運動の影響をうけるという問題点があった13).そこでこの実験においては,マウスに5~9秒間にわたり継続する音の刺激をあたえ,刺激の開始ではなく終了のタイミングを報告した場合に報酬をあたえた.このタスクにおいては音の刺激のあいだマウスは静止しているため,運動による影響をうけることなく音への応答を測定することが可能になった.
まず,1次聴覚野の2/3層の錐体細胞における神経活動をCa2+イメージング法により測定し,同一の細胞において,マウスがタスクに従事しているあいだの音への応答(行動時)と,受動的に音を聞いているあいだの音への応答(受動時)とを比較したところ,行動時においては錐体細胞の興奮性の応答が受動時よりも増加していた.この結果は,能動的な注意により知覚が向上するという心理学的な知見と一致するものであった.そこで,この可塑性の神経回路レベルでの基盤について明らかにするため,ふたたびトランスジェニックマウスを用いてCa2+イメージング法を細胞種に選択的に行った.その結果,1次聴覚野の4層の興奮性ニューロンにおける神経活動は行動時と受動時とで変化はなく,この可塑性が2/3層それ自体において誘導されていることが示された.パルブアルブミン陽性ニューロンおよびソマトスタチン陽性ニューロンにおいて同様に実験したところ,錐体細胞の場合とは逆に,ソマトスタチン陽性ニューロンの興奮性の応答のみが行動時において減少していた.これらの結果から,行動時においてはソマトスタチン陽性ニューロンにおける神経活動の低下が2/3層における錐体細胞の脱抑制をひき起こし,聴覚野における音への応答性を上昇させることが示唆された.
おわりに
この研究においては,マウスにおける長期にわたる生体内Ca2+イメージング法を用いて,受動的な音の経験のくり返しが1次聴覚野の2/3層における錐体細胞の音への応答性を日をおって低下させることを見い出した.逆に,マウスが音の刺激を用いたタスクに従事しているあいだは錐体細胞の音への応答性の一時的な上昇が観察された.したがって,1次聴覚野における音への応答は成熟した動物においても固定されたものではなく,音の刺激のもつ重要性に応じ双方向性に制御されることが明らかにされた.さらに,これらの双方向性の制御はいずれもソマトスタチン陽性ニューロンによる抑制の変化をともなっていた.これらの結果から,ソマトスタチン陽性ニューロンは感覚の刺激の重要性に応じ1次聴覚野における情報の流れを制御するゲートとしての役割をもつと考えられた(図2).
これまでの多くの研究において,成熟した動物の1次聴覚野における音への応答性は固定されたものであり,受動的な音の経験は可塑性を誘導しないと報告されてきた1,3,4).これらの報告とは異なり,この研究において可塑性が観察された理由として2つの可能性が考えられた.第1に,細胞外記録を用いた以前の電気生理学的な実験においては神経活動がどの細胞種に由来するのか不明であった点があげられる.この研究においては,2/3層の錐体細胞,4層の興奮性ニューロン,ソマトスタチン陽性ニューロンがそれぞれ異なる可塑性を示すことが明らかにされた.細胞外記録は高い神経活動を示す細胞からの記録にかたよる傾向が強いため,以前の研究においては,受動的な音の経験ののちにも馴化を起こさない4層の細胞やソマトスタチン陽性ニューロンから記録をとっていた可能性がある.第2に,この研究が覚醒したマウスを用いたのとは対照的に,以前の多くの実験は麻酔のもとであった点があげられる.脳は覚醒時と麻酔時において異なる神経活動のパターンを示すが,とくに,抑制性ニューロンの神経活動は麻酔のもとでは大きく低下することが脳の複数の部位において報告されている6,7).したがって,麻酔のもとでは抑制性のソマトスタチン陽性ニューロンの神経活動が低下し,結果として,可塑性の発現が抑えられた可能性が高い.
この研究では,1次聴覚野における局所的な神経回路に着目し,その可塑性におけるソマトスタチン陽性ニューロンの重要性を示した.それでは,ソマトスタチン陽性ニューロンそれ自体の神経活動はどのような機構により制御されているのだろうか.その可能性として,ノルアドレナリンやアセチルコリンなどの神経修飾物質,あるいは,前頭皮質など高次の領域からの入力といった,脳の領域をまたいだ相互作用の寄与が考えられる.これらの経路を光遺伝学などにより操作し,ソマトスタチン陽性ニューロンの神経活動,さらに,1次聴覚野の可塑性にどのような影響がみられるかを検討することが今後の課題である.将来的には,これまでの局所的な神経回路の研究から1歩進めて,局所の神経回路どうしをつなぐより大きなネットワークを遺伝学的なツールを用いて切り分けることにより,脳がいかにして状況に応じた情報処理をするのか,より包括的な理解を進めたい.
文 献
- Zhang, L. I., Bao, S. & Merzenich, M. M.: Persistent and specific influences of early acoustic environments on primary auditory cortex. Nat. Neurosci., 4, 1123-1130 (2001)[PubMed]
- De Villers-Sidani, E., Chang, E. F., Bao, S. et al.: Critical period window for spectral tuning defined in the primary auditory cortex (A1) in the rat. J. Neurosci., 27, 180-189 (2007)[PubMed]
- Bao, S., Chan, V. T. & Merzenich, M. M.: Cortical remodelling induced by activity of ventral tegmental dopamine neurons. Nature, 412, 79-83 (2001)[PubMed]
- Kilgard, M. P. & Merzenich, M. M.: Cortical map reorganization enabled by nucleus basalis activity. Science, 279, 1714-1718 (1998)[PubMed]
- Vallet, M., Maurin, M., Page, M. A. et al.: Annoyance from and habituation to road traffic noise from urban expressways. J. Sound Vib., 60, 423-440 (1978)
- Kato, H. K., Chu, M. W., Isaacson, J. S. et al.: Dynamic sensory representations in the olfactory bulb: modulation by wakefulness and experience. Neuron, 76, 962-975 (2012)[PubMed]
- Haider, B., Hausser, M. & Carandini, M.: Inhibition dominates sensory responses in the awake cortex. Nature, 493, 97-100 (2013)[PubMed]
- Isaacson, J. S. & Scanziani, M.: How inhibition shapes cortical activity. Neuron, 72, 231-243 (2011)[PubMed]
- Bregman, A. S.: Auditory Scene Analysis: The Perceptual Organization of Sound. MIT Press, Cambridge (1990)
- Fritz, J., Shamma, S., Elhilali, M. et al.: Rapid task-related plasticity of spectrotemporal receptive fields in primary auditory cortex. Nat. Neurosci., 6, 1216-1223 (2003)[PubMed]
- Rodgers, C. C. & DeWeese, M. R.: Neural correlates of task switching in prefrontal cortex and primary auditory cortex in a novel stimulus selection task for rodents. Neuron, 82, 1157-1170 (2014)[PubMed]
- David, S. V, Fritz, J. B. & Shamma, S. A.: Task reward structure shapes rapid receptive field plasticity in auditory cortex. Proc. Natl. Acad. Sci. USA, 109, 2144-2149 (2012)[PubMed]
- Schneider, D. M., Nelson, A. & Mooney, R.: A synaptic and circuit basis for corollary discharge in the auditory cortex. Nature, 513, 189-194 (2014)[PubMed]
著者プロフィール
略歴:2008年 東京大学大学院医学系研究科博士課程 修了,同年 同 博士研究員を経て,2010年より米国California大学San Diego校 博士研究員.
研究テーマ:生体における2光子励起イメージング法を用いた聴覚の情報処理の機構の解析.
Jeffry S. Isaacson
米国California大学San Diego校Professor.
研究室URL:http://isaacsonlab.net/Home.html
© 2015 加藤紘之・Jeffry S. Isaacson Licensed under CC 表示 2.1 日本
