方位選択性の神経活動に非依存的な初期形成および神経活動に依存した最適化
萩原賢太1・田川義晃2・大木研一1
(1九州大学大学院医学研究院 分子生理学分野,2京都大学大学院理学研究科 生物物理学系神経生物学分科)
email:萩原賢太,田川義晃,大木研一
DOI: 10.7875/first.author.2015.124
Neuronal activity is not required for the initial formation and maturation of visual selectivity.
Kenta M. Hagihara, Tomonari Murakami, Takashi Yoshida, Yoshiaki Tagawa, Kenichi Ohki
Nature Neuroscience, 18, 1780-1788 (2015)
大脳皮質の視覚野のニューロンは視覚入力の特定の傾き成分にのみ反応する方位選択性とよばれる性質を示す.この方位選択性は視覚情報の処理の根幹をなすと考えられており,その発見より研究が進められてきたが,発達期における形成の機構については依然としてよくわかっていない.今回,筆者らは,方位選択性の形成における神経活動の役割について調べるため,発達期において時期特異的に神経活動を抑制する技術と,マウスの視覚野における2光子Ca2+イメージング法とを組み合わせることにより,発達期において神経活動を抑制されたニューロンの反応の性質について調べた.その結果,方位選択性の初期形成は神経活動に依存しないが,ひきつづき起こる最適方位の最適化は神経活動に依存することが明らかにされた.この結果は,大脳皮質の機能の発達における自発的な神経活動の重要性を示すとともに,その初期形成において神経活動に依存しないどのような機構がかかわるのかという新たな問題を提起した.
方位選択性はその発見より1),性質それ自体が詳細に調べられるとともに,大脳における情報処理の機構の発達を調べるよいモデルとして用いられてきた.たとえば,縦縞の模様のみが見える環境で生育させた動物は横の方位の傾きを検知することができなくなることが知られており,このとき,正常に生育させた個体に比べ,視覚野において横の方位を最適方位とするニューロンの数が顕著に減少していることが示されている2).これは,方位の認識能という個体レベルでの情報処理の機構と,脳における情報表現がともに生後の経験により変化しうることを顕著に示している.しかし,方位選択性という反応の性質が生後どのように形成されるのかについては依然としてよくわかっていない.
網膜から外側膝状体,外側膝状体から大脳皮質へといたる視覚系神経回路3) や大脳皮質における神経回路4) の形成において神経活動が重要な役割を担うことから,方位選択性の形成にも神経活動がかかわるのではないかと考えられてきた.脳が未発達の状態で出生するフェレットの視覚野にテトロドトキシンを投与することにより方位選択性の形成が部分的に阻害されることが示されたが5),方位選択性の初期形成が起こると考えられる時期よりもまえからテトロドトキシンを継時的に投与することは技術的にむずかしく,初期形成における神経活動の役割は不明であった.また,すべてのニューロンの神経活動を阻害するテトロドトキシンの性質により,これまで,細胞自律的な神経活動の役割を調べることはできなかった.
方位選択性それ自体の形成の機構にくわえ,おのおののニューロンの最適方位がどのように決まっているのかも同様に大きな謎であった.以前に,筆者らのグループおよびほかのグループにより,同一の神経幹細胞に由来するニューロンは似た最適方位を示すことが明らかにされたが6,7),同一の神経幹細胞に由来するニューロンであっても異なる最適方位を示すニューロンも一定数が存在したことから,細胞分化ののちに最適方位が変わりうる可能性が示唆されており,その過程にもニューロンそれ自体の神経活動がかかわると考えられていた.
今回,これらの疑問に答えるべく,同一の個体において一部のニューロンの神経活動を時期特異的に抑制する技術と,生きた動物において個々のニューロンのレベルで神経活動を記録することのできる2光子Ca2+イメージング法8) とを組み合わせ,方位選択性の初期形成および最適方位の最適化の神経活動への依存性について調べた.
マウスの大脳皮質の視覚野において一部のニューロンの神経活動を抑制するために,内向き整流性K+チャネルであるKir2.1を子宮内電気穿孔法により1次視覚野の2/3層に導入する方法を用いた4).Kir2.1の時期特異的な発現を可能にするため,その発現をドキシサイクリンにより抑制するTet-off系を用いた.このとき,どのニューロンがKir2.1を発現するかを2光子イメージング法により同定するため,赤色蛍光タンパク質FP635を同時に導入した.
この神経活動の抑制手法の効果を確かめるため,開眼の前後の大脳皮質の自発的な神経活動,および,開眼ののちの視覚応答を2光子Ca2+イメージング法により調べたところ,Kir2.1を発現するニューロンおよび発現しないニューロンともに,自発的な神経活動は対照となるマウスと比べ顕著に減少していることがわかった.視覚応答に関しては,Kir2.1を発現するニューロンは視覚応答を示さず,Kir2.1を発現しないニューロンは正常な視覚応答を示した.
発達期において自発的な神経活動および視覚応答をともに抑制されたKir2.1発現ニューロンが方位選択性を正常に獲得したかどうかを調べるには,Kir2.1の発現をオフにして視覚応答を回復させる必要がある.そこで,Ca2+イメージング実験の1週間まえからドキシサイクリンの投与を開始し,Ca2+イメージング実験の際にはKir2.1の発現が十分に下がった状態になるようにした.すると,Kir2.1により発達期において神経活動を抑制されていたニューロンも正常な方位選択性を獲得していることがわかった.また,生後まもない段階から暗闇において生育させたマウスにおいても方位選択性は正常に形成されていた.すなわち,方位選択性の形成は自発的な神経活動および視覚経験のいずれにも依存しないことが明らかにされた.
正常に生育させた個体において視覚応答が開眼の前後にどのように変化するかについて調べた.開眼の直前の個体においては1次視覚野のニューロンは視覚応答をほとんど示さなかったが,自然な開眼の直後の個体においては成体と同じ程度のニューロンが視覚刺激に反応し,方位選択性を示すニューロンの割合も成体と同じ程度であった.このことから,視覚応答は開眼と同じ時期に急速に獲得され,方位選択性を可能にする神経回路は遅くとも開眼の直後には形成されていることがわかった.
最適方位が神経活動に依存して決定しているかどうかについて調べた.正常に成熟したマウスにおいては,縦向きに反応するニューロンおよび横向きに反応するニューロンが斜め方位に反応するニューロンに比べやや多く存在することが知られている.これは,自然界における視覚情報の方位成分の分布とよく一致することから,まわりの環境に対する脳における情報表現が最適化された結果であると考えられている9).まず,この最適化された分布が開眼の直後にすでに形成されているかどうか,正常の個体において調べた.すると,開眼の直後には横向きに反応するニューロンが縦向きに反応するニューロンよりも有意に多く,そののちの発達の過程において表現が最適化される,つまり,横向きに反応するニューロンと縦向きに反応するニューロンがほぼ同数になることがわかった.この過程については,開眼ののち,個々のニューロンが最適方位を変化させる,あるいは,視覚応答を示していなかったニューロンが縦向きを最適方位とした視覚応答を獲得する,という大きく2つの可能性が考えられたが,開眼の直後においても視覚応答を示すニューロンの割合は成熟した個体における割合と変わらなかったことから,個々のニューロンの最適方位が変化した可能性が高いと考えられた.
この最適方位の分布の最適化の過程が神経活動に依存的であるかどうかを検証するため,Kir2.1の発現あるいは暗室における飼育により神経活動を抑制したところ,暗室において飼育したマウスにおいて最適化は正常に起こるが,Kir2.1により神経活動を抑制されて生育したマウスにおいてはKir2.1を発現するニューロンおよび発現しないニューロンともに最適化が阻害されていた.さきに示したように,Kir2.1を導入したマウスのKir2.1発現ニューロンおよびKir2.1非発現ニューロンは,ともに開眼の前後において自発的な神経活動が抑制された.これらをあわせて考えると,最適方位の分布の最適化においては,視覚活動ではなく自発的な神経活動が重要であることが示唆された.
以上の一連の実験により,方位選択性の初期形成は神経活動に非依存的に起こり,開眼の直後には視覚応答はすでに成体と同じ程度まで発達しているが,その時点では最適方位の分布は最適化されておらず,のちの自発的な神経活動により最適化が起こる,という2段階の発達の様式を示すことが明らかにされた(図1).
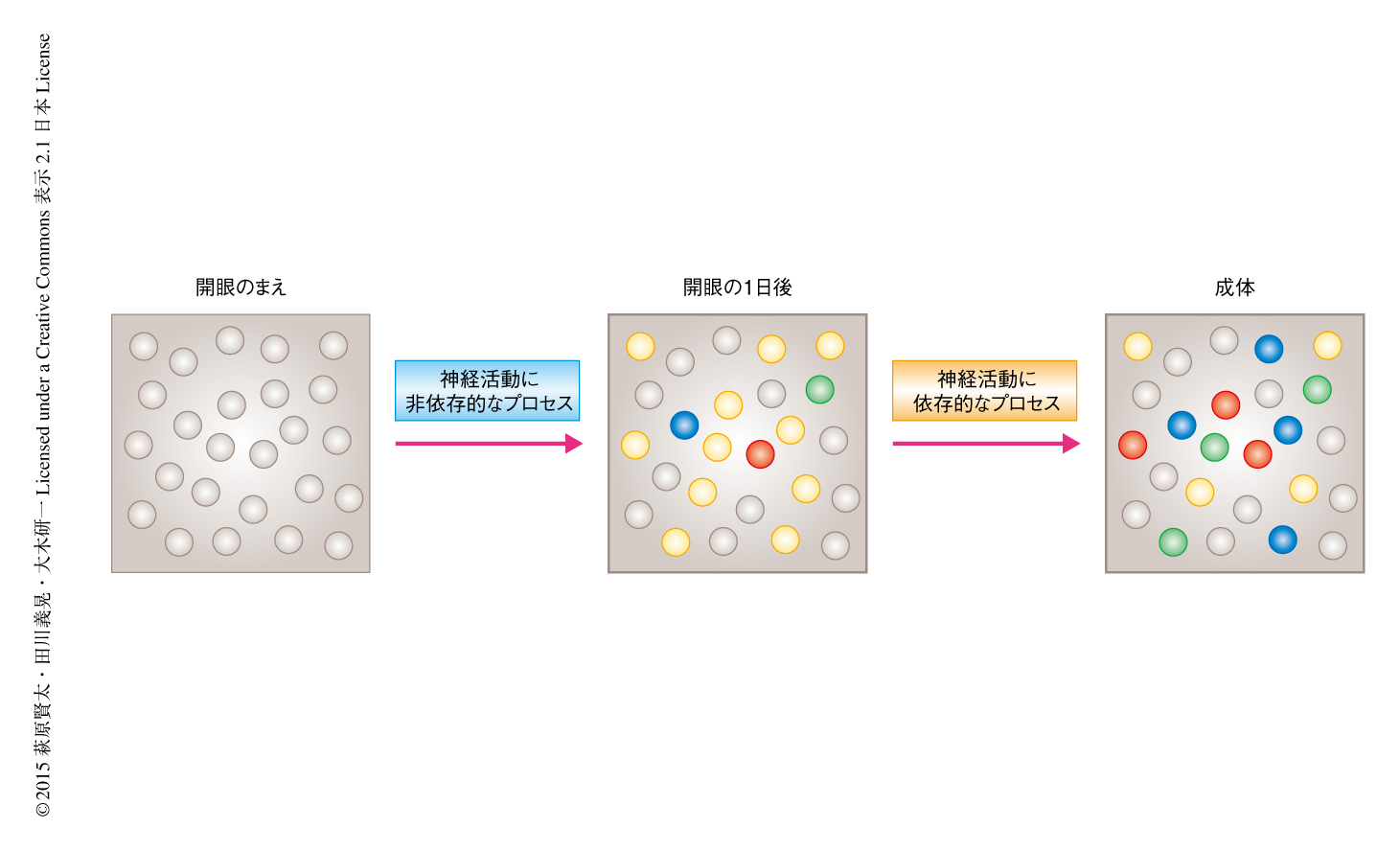
この研究により,方位選択性の初期形成は神経活動に非依存的に起こることがわかったが,ではいったい何がこれを可能にしているのであろうか.さきに述べたように,最適方位の決定においてはどの神経幹細胞から分化するかという細胞系譜が重要なことが示されているが6,7),同時に,このとき,同一の神経幹細胞に由来するニューロンにおいてギャップ結合の作用を阻害すると,似た最適方位の獲得が起こらないことも示されている7).方位選択性それ自体の初期形成においても,特定の分子が重要なはたらきをもち,ある程度までハードワイヤードに形成される可能性は十分にあると考えられる.
今回,視覚皮質における神経活動が2/3層および4層がともに抑制された状態においても方位選択性は正常に形成されることが示されたが,この実験では視覚視床における神経活動は抑制されていない.そのため,外側膝状体をはじめとする視覚視床における神経活動に由来する,視覚野のニューロンの樹状突起における活動電位をひき起こす閾値にみたない神経活動だけで方位選択性が形成される可能性は完全には否定できない.また,これまで視覚視床からの入力はほとんどが方位選択性を示さないと考えられていたが,近年,方向選択性を示す網膜のニューロンからの入力が外側膝状体の外側にあるニューロンを介して1次視覚野に到達していることが報告されており10),これらの選択性をもつ入力が1次視覚野における方位選択性の初期形成にどのように寄与しているかについても興味深い.
開眼の直後の横向きに反応するニューロンが多い状態から,自発的な神経活動により横向きに反応するニューロンと縦向きに反応するニューロンがほぼ均等に分布する状態になる機構も今後の検討課題である.自発的な神経活動は大脳皮質の局所における神経回路の結合の様式をどのように制御することにより,この最適方位の最適化を誘導するのであろうか.また,その機構は視覚経験に依存的な最適方位の変化2) を可能にする機構と共通なのであろうか.なお,今回,提唱した最適方位の最適化のモデル(図1)は,あくまで開眼の直後と成長ののちの点での観察にもとづく推測であり,さらなる検証が必要と考えられる.高感度の遺伝的なCa2+指示タンパク質を用いた継時的なイメージング法により11,12),この最適化の過程を直接的に観察することがつぎの重要なステップになるだろう.
略歴:九州大学大学院医学研究院博士課程 在学中.
研究テーマ:情報処理,記憶,学習を可能にする神経可塑性をひき起こす神経活動.
抱負:神経回路のレベルからボトムアップに意識の解明にむけて研究をつづけたい.
田川 義晃(Yoshiaki Tagawa)
京都大学大学院理学研究科 講師.
大木 研一(Kenichi Ohki)
九州大学大学院医学研究院 教授.
研究室URL:http://www.physiol2.med.kyushu-u.ac.jp/
© 2015 萩原賢太・田川義晃・大木研一 Licensed under CC 表示 2.1 日本
(1九州大学大学院医学研究院 分子生理学分野,2京都大学大学院理学研究科 生物物理学系神経生物学分科)
email:萩原賢太,田川義晃,大木研一
DOI: 10.7875/first.author.2015.124
Neuronal activity is not required for the initial formation and maturation of visual selectivity.
Kenta M. Hagihara, Tomonari Murakami, Takashi Yoshida, Yoshiaki Tagawa, Kenichi Ohki
Nature Neuroscience, 18, 1780-1788 (2015)
要 約
大脳皮質の視覚野のニューロンは視覚入力の特定の傾き成分にのみ反応する方位選択性とよばれる性質を示す.この方位選択性は視覚情報の処理の根幹をなすと考えられており,その発見より研究が進められてきたが,発達期における形成の機構については依然としてよくわかっていない.今回,筆者らは,方位選択性の形成における神経活動の役割について調べるため,発達期において時期特異的に神経活動を抑制する技術と,マウスの視覚野における2光子Ca2+イメージング法とを組み合わせることにより,発達期において神経活動を抑制されたニューロンの反応の性質について調べた.その結果,方位選択性の初期形成は神経活動に依存しないが,ひきつづき起こる最適方位の最適化は神経活動に依存することが明らかにされた.この結果は,大脳皮質の機能の発達における自発的な神経活動の重要性を示すとともに,その初期形成において神経活動に依存しないどのような機構がかかわるのかという新たな問題を提起した.
はじめに
方位選択性はその発見より1),性質それ自体が詳細に調べられるとともに,大脳における情報処理の機構の発達を調べるよいモデルとして用いられてきた.たとえば,縦縞の模様のみが見える環境で生育させた動物は横の方位の傾きを検知することができなくなることが知られており,このとき,正常に生育させた個体に比べ,視覚野において横の方位を最適方位とするニューロンの数が顕著に減少していることが示されている2).これは,方位の認識能という個体レベルでの情報処理の機構と,脳における情報表現がともに生後の経験により変化しうることを顕著に示している.しかし,方位選択性という反応の性質が生後どのように形成されるのかについては依然としてよくわかっていない.
網膜から外側膝状体,外側膝状体から大脳皮質へといたる視覚系神経回路3) や大脳皮質における神経回路4) の形成において神経活動が重要な役割を担うことから,方位選択性の形成にも神経活動がかかわるのではないかと考えられてきた.脳が未発達の状態で出生するフェレットの視覚野にテトロドトキシンを投与することにより方位選択性の形成が部分的に阻害されることが示されたが5),方位選択性の初期形成が起こると考えられる時期よりもまえからテトロドトキシンを継時的に投与することは技術的にむずかしく,初期形成における神経活動の役割は不明であった.また,すべてのニューロンの神経活動を阻害するテトロドトキシンの性質により,これまで,細胞自律的な神経活動の役割を調べることはできなかった.
方位選択性それ自体の形成の機構にくわえ,おのおののニューロンの最適方位がどのように決まっているのかも同様に大きな謎であった.以前に,筆者らのグループおよびほかのグループにより,同一の神経幹細胞に由来するニューロンは似た最適方位を示すことが明らかにされたが6,7),同一の神経幹細胞に由来するニューロンであっても異なる最適方位を示すニューロンも一定数が存在したことから,細胞分化ののちに最適方位が変わりうる可能性が示唆されており,その過程にもニューロンそれ自体の神経活動がかかわると考えられていた.
今回,これらの疑問に答えるべく,同一の個体において一部のニューロンの神経活動を時期特異的に抑制する技術と,生きた動物において個々のニューロンのレベルで神経活動を記録することのできる2光子Ca2+イメージング法8) とを組み合わせ,方位選択性の初期形成および最適方位の最適化の神経活動への依存性について調べた.
1.時期特異的な神経活動の抑制
マウスの大脳皮質の視覚野において一部のニューロンの神経活動を抑制するために,内向き整流性K+チャネルであるKir2.1を子宮内電気穿孔法により1次視覚野の2/3層に導入する方法を用いた4).Kir2.1の時期特異的な発現を可能にするため,その発現をドキシサイクリンにより抑制するTet-off系を用いた.このとき,どのニューロンがKir2.1を発現するかを2光子イメージング法により同定するため,赤色蛍光タンパク質FP635を同時に導入した.
この神経活動の抑制手法の効果を確かめるため,開眼の前後の大脳皮質の自発的な神経活動,および,開眼ののちの視覚応答を2光子Ca2+イメージング法により調べたところ,Kir2.1を発現するニューロンおよび発現しないニューロンともに,自発的な神経活動は対照となるマウスと比べ顕著に減少していることがわかった.視覚応答に関しては,Kir2.1を発現するニューロンは視覚応答を示さず,Kir2.1を発現しないニューロンは正常な視覚応答を示した.
2.神経活動に非依存的な方位選択性の初期形成
発達期において自発的な神経活動および視覚応答をともに抑制されたKir2.1発現ニューロンが方位選択性を正常に獲得したかどうかを調べるには,Kir2.1の発現をオフにして視覚応答を回復させる必要がある.そこで,Ca2+イメージング実験の1週間まえからドキシサイクリンの投与を開始し,Ca2+イメージング実験の際にはKir2.1の発現が十分に下がった状態になるようにした.すると,Kir2.1により発達期において神経活動を抑制されていたニューロンも正常な方位選択性を獲得していることがわかった.また,生後まもない段階から暗闇において生育させたマウスにおいても方位選択性は正常に形成されていた.すなわち,方位選択性の形成は自発的な神経活動および視覚経験のいずれにも依存しないことが明らかにされた.
3.開眼の前後における視覚反応の獲得
正常に生育させた個体において視覚応答が開眼の前後にどのように変化するかについて調べた.開眼の直前の個体においては1次視覚野のニューロンは視覚応答をほとんど示さなかったが,自然な開眼の直後の個体においては成体と同じ程度のニューロンが視覚刺激に反応し,方位選択性を示すニューロンの割合も成体と同じ程度であった.このことから,視覚応答は開眼と同じ時期に急速に獲得され,方位選択性を可能にする神経回路は遅くとも開眼の直後には形成されていることがわかった.
4.神経活動に依存的な最適方位の最適化
最適方位が神経活動に依存して決定しているかどうかについて調べた.正常に成熟したマウスにおいては,縦向きに反応するニューロンおよび横向きに反応するニューロンが斜め方位に反応するニューロンに比べやや多く存在することが知られている.これは,自然界における視覚情報の方位成分の分布とよく一致することから,まわりの環境に対する脳における情報表現が最適化された結果であると考えられている9).まず,この最適化された分布が開眼の直後にすでに形成されているかどうか,正常の個体において調べた.すると,開眼の直後には横向きに反応するニューロンが縦向きに反応するニューロンよりも有意に多く,そののちの発達の過程において表現が最適化される,つまり,横向きに反応するニューロンと縦向きに反応するニューロンがほぼ同数になることがわかった.この過程については,開眼ののち,個々のニューロンが最適方位を変化させる,あるいは,視覚応答を示していなかったニューロンが縦向きを最適方位とした視覚応答を獲得する,という大きく2つの可能性が考えられたが,開眼の直後においても視覚応答を示すニューロンの割合は成熟した個体における割合と変わらなかったことから,個々のニューロンの最適方位が変化した可能性が高いと考えられた.
この最適方位の分布の最適化の過程が神経活動に依存的であるかどうかを検証するため,Kir2.1の発現あるいは暗室における飼育により神経活動を抑制したところ,暗室において飼育したマウスにおいて最適化は正常に起こるが,Kir2.1により神経活動を抑制されて生育したマウスにおいてはKir2.1を発現するニューロンおよび発現しないニューロンともに最適化が阻害されていた.さきに示したように,Kir2.1を導入したマウスのKir2.1発現ニューロンおよびKir2.1非発現ニューロンは,ともに開眼の前後において自発的な神経活動が抑制された.これらをあわせて考えると,最適方位の分布の最適化においては,視覚活動ではなく自発的な神経活動が重要であることが示唆された.
以上の一連の実験により,方位選択性の初期形成は神経活動に非依存的に起こり,開眼の直後には視覚応答はすでに成体と同じ程度まで発達しているが,その時点では最適方位の分布は最適化されておらず,のちの自発的な神経活動により最適化が起こる,という2段階の発達の様式を示すことが明らかにされた(図1).
おわりに
この研究により,方位選択性の初期形成は神経活動に非依存的に起こることがわかったが,ではいったい何がこれを可能にしているのであろうか.さきに述べたように,最適方位の決定においてはどの神経幹細胞から分化するかという細胞系譜が重要なことが示されているが6,7),同時に,このとき,同一の神経幹細胞に由来するニューロンにおいてギャップ結合の作用を阻害すると,似た最適方位の獲得が起こらないことも示されている7).方位選択性それ自体の初期形成においても,特定の分子が重要なはたらきをもち,ある程度までハードワイヤードに形成される可能性は十分にあると考えられる.
今回,視覚皮質における神経活動が2/3層および4層がともに抑制された状態においても方位選択性は正常に形成されることが示されたが,この実験では視覚視床における神経活動は抑制されていない.そのため,外側膝状体をはじめとする視覚視床における神経活動に由来する,視覚野のニューロンの樹状突起における活動電位をひき起こす閾値にみたない神経活動だけで方位選択性が形成される可能性は完全には否定できない.また,これまで視覚視床からの入力はほとんどが方位選択性を示さないと考えられていたが,近年,方向選択性を示す網膜のニューロンからの入力が外側膝状体の外側にあるニューロンを介して1次視覚野に到達していることが報告されており10),これらの選択性をもつ入力が1次視覚野における方位選択性の初期形成にどのように寄与しているかについても興味深い.
開眼の直後の横向きに反応するニューロンが多い状態から,自発的な神経活動により横向きに反応するニューロンと縦向きに反応するニューロンがほぼ均等に分布する状態になる機構も今後の検討課題である.自発的な神経活動は大脳皮質の局所における神経回路の結合の様式をどのように制御することにより,この最適方位の最適化を誘導するのであろうか.また,その機構は視覚経験に依存的な最適方位の変化2) を可能にする機構と共通なのであろうか.なお,今回,提唱した最適方位の最適化のモデル(図1)は,あくまで開眼の直後と成長ののちの点での観察にもとづく推測であり,さらなる検証が必要と考えられる.高感度の遺伝的なCa2+指示タンパク質を用いた継時的なイメージング法により11,12),この最適化の過程を直接的に観察することがつぎの重要なステップになるだろう.
文 献
- Hubel, D. H. & Wiesel, T. N.: Receptive fields of single neurons in the cat’s striate cortex. J. Physiol., 148, 574-591 (1959)[PubMed]
- Blakemore, C. & Cooper, C. F.: Development of the brain depends on the visual environment. Nature, 228, 477-478 (1970)[PubMed]
- Huberman, A. D., Feller, M. B. & Chapman, B.: Mechanisms underlying development of visual maps and receptive fields. Annu. Rev. Neurosci., 31, 479-509 (2008)[PubMed]
- Mizuno, H., Hirano, T. & Tagawa, Y.: Evidence for activity-dependent cortical wiring: formation of interhemispheric connections in neonatal mouse visual cortex requires projection neuron activity. J. Neurosci., 27, 6760-6770 (2007)[PubMed]
- Chapman, B. & Stryker, M. P.: Development of orientation selectivity in ferret visual cortex and effects of deprivation. J. Neurosci., 13, 5251-5262 (1993)[PubMed]
- Ohtsuki, G., Nishiyama, M., Yoshida, T. et al.: Similarity of visual selectivity among clonally related neurons in visual cortex. Neuron, 75, 65-72 (2012)[PubMed]
- Li, Y., Lu, H., Cheng, P. L. et al.: Clonally related visual cortical neurons show similar stimulus feature selectivity. Nature, 486, 118-121 (2012)[PubMed]
- Ohki, K., Chung, S., Ch'ng, Y. H. et al.: Functional imaging with cellular resolution reveals precise micro-architecture in visual cortex. Nature, 433, 597-603 (2005)[PubMed]
- Coppola, D. M., White, L. E., Fitzpatrick, D. et al.: Unequal representation of cardinal and oblique contours in ferret visual cortex. Proc. Natl. Acad. Sci. USA, 95, 2621-2623 (1998)[PubMed]
- Cruz-Martin, A., El-Danaf, R. N., Osakada, F. et al.: A dedicated circuit links direction-selective retinal ganglion cells to the primary visual cortex. Nature, 507, 358-361 (2014)[PubMed]
- Ohkura, M., Sasaki, T., Sadakari, J. et al.: Genetically encoded green fluorescent Ca2+ indicators with improved detectability for neuronal Ca2+ signals. PLoS One, 7, e51286 (2012)[PubMed]
- Chen, T. W., Wardill, T. J., Sun, Y. et al.: Ultrasensitive fluorescent proteins for imaging neuronal activity. Nature, 499, 295-300 (2013)[PubMed]
著者プロフィール
略歴:九州大学大学院医学研究院博士課程 在学中.
研究テーマ:情報処理,記憶,学習を可能にする神経可塑性をひき起こす神経活動.
抱負:神経回路のレベルからボトムアップに意識の解明にむけて研究をつづけたい.
田川 義晃(Yoshiaki Tagawa)
京都大学大学院理学研究科 講師.
大木 研一(Kenichi Ohki)
九州大学大学院医学研究院 教授.
研究室URL:http://www.physiol2.med.kyushu-u.ac.jp/
© 2015 萩原賢太・田川義晃・大木研一 Licensed under CC 表示 2.1 日本
