マウスの大脳皮質において知覚の形成に必須なトップダウンの制御
真仁田 聡・鈴木崇之・村山正宜
(理化学研究所脳科学総合研究センター 行動神経生理学研究チーム)
email:真仁田 聡,鈴木崇之,村山正宜
DOI: 10.7875/first.author.2015.076
A top-down cortical circuit for accurate sensory perception.
Satoshi Manita, Takayuki Suzuki, Chihiro Homma, Takashi Matsumoto, Maya Odagawa, Kazuyuki Yamada, Keisuke Ota, Chie Matsubara, Ayumu Inutsuka, Masaaki Sato, Masamichi Ohkura, Akihiro Yamanaka, Yuchio Yanagawa, Junichi Nakai, Yasunori Hayashi, Matthew E. Larkum, Masanori Murayama
Neuron, 86, 1304-1316 (2015)
これまで,高次の脳領域から感覚野へのトップダウンの入力は知覚をつかさどる神経回路の活動を修飾していると考えられてきた.この研究において,マウスの体性感覚野に対するトップダウンの神経回路の解剖学的および機能的な特徴が明らかにされ,トップダウンの入力が正確な知覚の形成に必須であることが示された.皮膚に存在する感覚器からの情報は,視床などを介しボトムアップの入力として大脳皮質の第1体性感覚野を活性化するが,この第1体性感覚野の活動はより高次な脳領域である第2運動野へと伝達され,活性化された第2運動野はふたたび第1体性感覚野を活性化するという反響回路が見い出された.この神経回路において,第2運動野からのトップダウンの入力は第1体性感覚野の深層および表層に集中し,第5層の錐体細胞の樹状突起においてCa2+スパイクをともなう持続的な発火活動を誘発した.さらに,このトップダウンの入力を抑制すると第5層の錐体細胞におけるこれらの神経活動が抑制され,マウスは正常な知覚行動ができなくなった.これらの結果から,正常な知覚の形成にはトップダウンの入力が必須であることが示された.
皮膚においてうけとる外界からの情報は脊髄や視床を経由して第1体性感覚野に伝達され,さらにより高次な脳領域へボトムアップな情報として伝達される.このボトムアップの入力だけで知覚を形成できるのだろうか.われわれは高次の脳領域が関連する注意や予測といった情報が知覚に影響をあたえることを主観的に経験している.このことは,知覚の形成においてはボトムアップの入力だけでなくトップダウンの入力も関連することを示す.しかし,知覚をつかさどる神経回路に対するトップダウンの入力の役割は十分に検証されていない.げっ歯類において第1体性感覚野に対する高次の脳領域はいくつか存在し,そのひとつとして第2運動野がある.第2運動野は大脳皮質の前方に存在し,価値にもとづく意思決定1) や随意運動2) といった高次の脳機能との関連が示唆されてきた.第2運動野と第1体性感覚野とは解剖学的に互いに結合していることから,第1体性感覚野における神経活動は第2運動野からの入力により制御されていると考えられる.そこで,この研究においては,知覚に対するトップダウンの入力の影響を明らかにするため,第2運動野および第1体性感覚野から構成される神経回路の解剖学的および生理学的な同定を試みた.さらに,光遺伝学的な手法を用いた行動実験により,マウスの知覚行動に対する第2運動野から第1体性感覚野へのトップダウンの入力の役割について調べた.
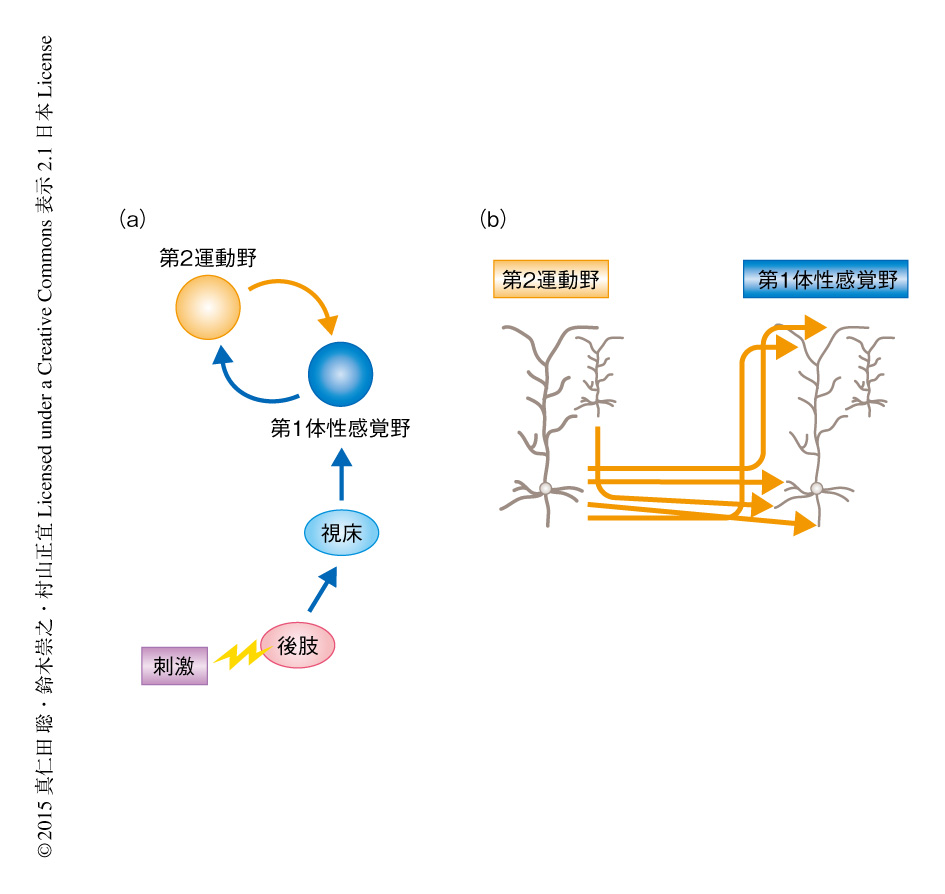
マウスの後肢への刺激により第1体性感覚野および第2運動野が活性化されるかどうか検討するため,膜電位の変化により蛍光強度の変化する色素を用いて膜電位イメージング実験を行った.大脳皮質の広範囲な領域から神経活動を記録したところ,後肢への刺激によりまず第1体性感覚野が活動し,それにひきつづき第2運動野が活動することが見い出された.第1体性感覚野と第2運動野とが機能的に相互に連絡しているかどうか検討するため,それぞれの部位における神経活動を薬理学的に抑制した.その結果,第1体性感覚野を抑制したときは第2運動野における神経活動において早い成分が,第2運動野を抑制したときは第1体性感覚野における神経活動において遅い成分が,それぞれ減少した.これらの結果から,まず,後肢からのボトムアップの入力が第1体性感覚野を活性化し,ついで,第1体性感覚野が第2運動野を活性化し,そして,第2運動野がふたたび第1体性感覚野を活性化する,という反響回路の存在が示された(図1a).この反響回路の解剖学的な特徴を調べるため,第1体性感覚野あるいは第2運動野に順行性の神経トレーサーを注入した.その結果,第1体性感覚野から第2運動野へ,および,第2運動野から第1体性感覚野へ,それぞれ投射する神経軸索が確認された.第2運動野から第1体性感覚野へ投射する神経軸索は深層および表層において密であった(図1b).この投射パターンは典型的なトップダウンの入力の投射パターンと同様であった3).
神経トレーサーを用いた実験から,第2運動野からのトップダウンの入力は第1体性感覚野の深層および表層を活性化すると考えられた.これを検討するため,16点の記録部位が一列にならんだ電極によるマルチユニット記録法を用いて,第1体性感覚野のすべての層(第1層~第6層)から活動電位の応答を記録した.この手法により,1つの記録部位から複数の単一のニューロンの活動電位を記録することができる.後肢への刺激はすべての層において早発性の神経活動および遅発性の神経活動を誘起した.早発性の神経活動は第4層においてもっとも早くピークに達した.これは視床からのボトムアップの入力により誘起された神経活動であると考えられた.一方,遅発性の神経活動は深層から表層という順に生じた.膜電位イメージング実験において,第1体性感覚野における神経活動において遅い成分は第2運動野の神経活動に依存していたことから,この遅発性の神経活動は第2運動野における神経活動により誘起されると考えられた.これを検証するため,第2運動野における神経活動を薬理学的に抑制したところ,遅発性の神経活動だけが抑制された.この結果から,遅発性の神経活動が第2運動野からのトップダウンの入力に起因することが示された.遅発性の神経活動において第5層における神経活動の大きさはどの層よりも顕著であったが,これは第2運動野から第1体性感覚野への神経軸索の投射パターンから予測される神経活動とは一致しなかった.
第2運動野からの神経軸索は第1体性感覚野の深層および表層に投射していたにもかかわらず,第2運動野における神経活動に由来する遅発性の神経活動は第5層において顕著であった.この理由のひとつとして,第2運動野から第1体性感覚野へのシナプス入力は第5層に集中している可能性が考えられた.そこで,第2運動野から第1体性感覚野への興奮性のシナプス入力の強度を推定するため,第1体性感覚野において記録した細胞外電位から電流源密度解析を行った.その結果,マルチユニット記録法により得られた結果と同じように,後肢への刺激によりすべての層において早発性のシナプス入力および遅発性のシナプス入力が生じた.遅発性のシナプス入力の大きさは第1体性感覚野の深層および表層において顕著であり,これは第2運動野からの投射パターンと一致していた.
第1体性感覚野の深層および表層におけるシナプス入力はおよそ30ミリ秒のあいだ重複していた.第5層の錐体細胞は表層まで樹状突起を伸長していることを考えると,この細胞は30ミリ秒のあいだに深層にある基底の樹状突起および表層にある遠位の樹状突起により第2運動野からの入力をうけ,これを統合している可能性が考えられた.これまでの研究により,第5層の錐体細胞が30ミリ秒のあいだにこれらの入力を統合することにより遠位の樹状突起においてCa2+スパイクの発生することが知られている4).また,このCa2+スパイクにより細胞体においては持続的な発火活動が誘起される5,6).そこで,神経トレーサー実験の結果とマルチユニット記録における結果の不一致は,第1体性感覚野の深層および表層へのシナプス入力が第5層の錐体細胞の樹状突起においてCa2+スパイクを誘起し,これにより持続的な発火活動がひき起こされたためであるとの仮説をたてた.これを検証するため,2光子イメージング法を用いて第5層の錐体細胞の遠位の樹状突起からのCa2+応答を記録したところ,後肢への刺激により“小さくて早いCa2+応答”および“大きくて遅いCa2+応答”が観察された.薬理学的な実験により,この大きくて遅いCa2+応答は第2運動野の神経活動に依存的であることが示され,また,このCa2+応答はCa2+スパイクであることが示唆された.この結果およびマルチユニット記録における薬理学的な実験の結果から,第2運動野からの入力が第5層の錐体細胞の樹状突起においてCa2+スパイクを誘起し,これにより細胞体において持続的な発火活動が生じたことが示された.
第2運動野からのトップダウンの入力がマウスの知覚行動にあたえる影響について調べるため,後肢への触覚にもとづく行動課題を行った.第2運動野から第1体性感覚野への神経軸索の投射を光遺伝学的に抑制するため,第2運動野のニューロンに光感受性のイオンポンプを発現させたマウスを用いた.まず,自発性の場所選好試験を行った.この実験では,箱のなかに2種類の異なる床面,ツルツル床およびザラザラ床を配置した.光を照射されないマウスはどちらか一方の床面に対し滞在時間のかたより(つまり,選好性)を示したが,光の照射により第2運動野から第1体性感覚野への入力を抑制したマウスはその滞在時間のかたよりが減少した.この結果は,光の照射により床面に対する知覚が変化したためと考えられた.つぎに,刺激誘発性の肢動作試験を行った.マウスは後肢を軽く刺激されるとある確率で肢を動かす.光の照射により第2運動野から第1体性感覚野への入力を抑制したとき,刺激された肢を動かす確率がどう変化するかを検討した.このとき,第1体性感覚野にて生じる第2運動野からの入力に由来する遅発性の神経活動を選択的に抑制した.その結果,光を照射しない条件と比べ肢を動かす確率は低下し,後肢への刺激により誘発される遅発性の神経活動が知覚の形成に必要であることが示された.
皮膚感覚の知覚が第2運動野から第1体性感覚野へのトップダウンの入力の抑制により変容しているなら,その知覚をもとにした高次機能を示す行動も変化する可能性が考えられた.これを検証するため,Y字型迷路による床面弁別行動試験を行った.この実験では,Y字型迷路の分岐点の手前にツルツル床あるいはザラザラ床を配置し,おのおのの触感に対応した方向に報酬を提示した(たとえば,ザラザラ床の場合には左の方向に報酬が提示される).マウスは訓練により床面の触感と報酬が提示される方向との関係を約80%の正解率まで学習した.これらのマウスを用い,光の照射により第2運動野から第1体性感覚野への入力を抑制すると,正解率は約65%まで低下した.一方,光の照射により,運動,視覚の能力,不安様の行動などに影響はみられなかった.これらの行動実験より,正常な知覚行動には第2運動野から第1体性感覚野へのトップダウンの入力が必須であることが明らかにされた.
この研究において,マウスの体性感覚において第2運動野から第1体性感覚野へのトップダウンの入力の神経回路およびその生理的な役割が明らかにされた.具体的には,1)視床からのボトムアップの入力により第1体性感覚野において早発性の神経活動が生じ,これが第2運動野へと伝達される.活性化された第2運動野は第1体性感覚野における遅発性の神経活動を誘起する.2)第2運動野からのトップダウンの入力は第1体性感覚野の深層および表層へと投射され,これらの入力により第5層の錐体細胞の樹状突起においてCa2+スパイクが発生し,細胞体において持続的な発火活動が生じる.3)第2運動野からの入力を第1体性感覚野において光遺伝学的に抑制するとマウスの正確な知覚行動が変化する.
これまでも,第1次感覚野において感覚刺激により誘起される遅発性の神経活動については報告されており,この神経活動の振幅と行動課題の成績とのあいだに強い正の相関のあることが示唆されていた7-11).しかし,この遅発性の神経活動が知覚に必須なのか,それとも,知覚の体験による余波としての神経活動なのかは明らかにされていなかった.最近の報告によると,マウスを用いたヒゲへの刺激の弁別課題において,第1体性感覚野における抑制性のニューロンを光遺伝学的に活性化することにより遅発性の神経活動を抑制すると,マウスはあたかもヒゲへの刺激を知覚していないような行動を示す10).この結果および今回の結果は,遅発性の神経活動は知覚に必須であることを示す.さらに今回,この遅発性の神経活動の実体は第2運動野からのトップダウンの入力による第5層の錐体細胞の樹状突起におけるCa2+スパイクとそれにつづく持続した発火活動であることが明らかにされた.
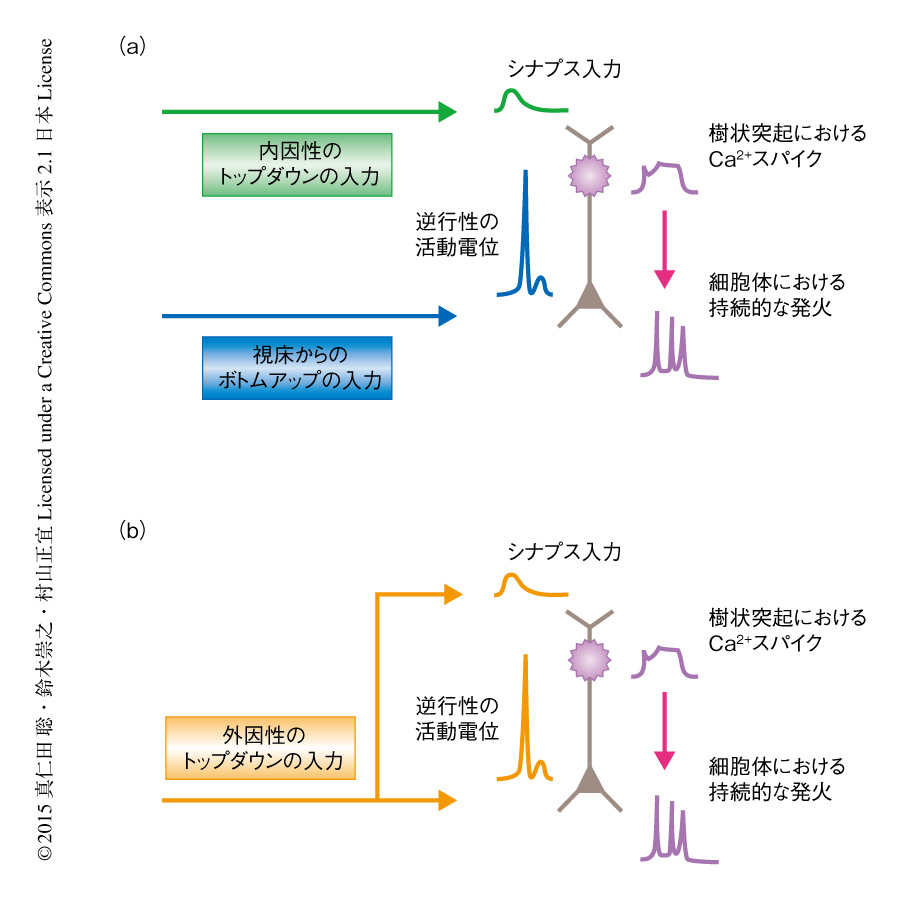
われわれはどのような神経活動により知覚を体験するのか.単一のニューロンのレベルでの知覚モデルのひとつとして,連合入力モデルがある(図2a).われわれが知覚するためには,視床に由来するボトムアップの入力とは別に,注意や予測といった脳内の活動による“内因性”のトップダウンの入力も必要とするというモデルである.ボトムアップの入力および内因性のトップダウンの入力が脳のある領域において短い時間のあいだ連合することにより樹状突起においてCa2+スパイクが発生し,それにつづく細胞体における持続的な発火活動が知覚をひき起こす.もしこの仮説が正しければ,われわれは“なにかに注意しなければなにも感じない”ことにもなる.しかし実際には,とくに注意せずぼんやりしている状態でも皮膚の感覚を知覚することは可能であり,この仮説ではわれわれの実体験をすべて説明することはできない.今回,見い出された反響回路では,肢からの情報が第1体性感覚野を経由して第2運動野において“外因性”のトップダウンの入力に変換され第1体性感覚野にフィードバックされる(図2b).筆者らは,従来の連合入力モデルと同様に,この外因性のトップダウンの入力のみで樹状突起においてCa2+スパイクをひき起こす機能を見い出した.この反響回路が,われわれがとくに注意などしていない状態においても知覚の形成を可能にする基本的な神経回路として動作している可能性がある.脳はそのときどきの状態により,連合入力モデルによる神経回路と反響回路とを使い分けていると考えられる.
略歴:2006年東京薬科大学大学院生命科学研究科 修了,同年 米国New York Medical College研究員,2010年 理化学研究所脳科学総合研究センター 研究員を経て,2015年より山梨大学医学部 特任助教.
研究テーマ:生きている動物の脳における単一のニューロンの特性.
抱負:脳切片標本から得られた知見が生きた脳においても適応できるのかといった問題や,単一のニューロンの活動と動物の行動や学習といった脳の高次機能との因果関係について明らかにしたい.
鈴木 崇之(Takayuki Suzuki)
理化学研究所脳科学総合研究センター 研究員.
村山 正宜(Masanori Murayama)
理化学研究所脳科学総合研究センター チームリーダー.
研究室URL:http://murayama-lab.brain.riken.jp/index_jp.html
© 2015 真仁田 聡・鈴木崇之・村山正宜 Licensed under CC 表示 2.1 日本
(理化学研究所脳科学総合研究センター 行動神経生理学研究チーム)
email:真仁田 聡,鈴木崇之,村山正宜
DOI: 10.7875/first.author.2015.076
A top-down cortical circuit for accurate sensory perception.
Satoshi Manita, Takayuki Suzuki, Chihiro Homma, Takashi Matsumoto, Maya Odagawa, Kazuyuki Yamada, Keisuke Ota, Chie Matsubara, Ayumu Inutsuka, Masaaki Sato, Masamichi Ohkura, Akihiro Yamanaka, Yuchio Yanagawa, Junichi Nakai, Yasunori Hayashi, Matthew E. Larkum, Masanori Murayama
Neuron, 86, 1304-1316 (2015)
要 約
これまで,高次の脳領域から感覚野へのトップダウンの入力は知覚をつかさどる神経回路の活動を修飾していると考えられてきた.この研究において,マウスの体性感覚野に対するトップダウンの神経回路の解剖学的および機能的な特徴が明らかにされ,トップダウンの入力が正確な知覚の形成に必須であることが示された.皮膚に存在する感覚器からの情報は,視床などを介しボトムアップの入力として大脳皮質の第1体性感覚野を活性化するが,この第1体性感覚野の活動はより高次な脳領域である第2運動野へと伝達され,活性化された第2運動野はふたたび第1体性感覚野を活性化するという反響回路が見い出された.この神経回路において,第2運動野からのトップダウンの入力は第1体性感覚野の深層および表層に集中し,第5層の錐体細胞の樹状突起においてCa2+スパイクをともなう持続的な発火活動を誘発した.さらに,このトップダウンの入力を抑制すると第5層の錐体細胞におけるこれらの神経活動が抑制され,マウスは正常な知覚行動ができなくなった.これらの結果から,正常な知覚の形成にはトップダウンの入力が必須であることが示された.
はじめに
皮膚においてうけとる外界からの情報は脊髄や視床を経由して第1体性感覚野に伝達され,さらにより高次な脳領域へボトムアップな情報として伝達される.このボトムアップの入力だけで知覚を形成できるのだろうか.われわれは高次の脳領域が関連する注意や予測といった情報が知覚に影響をあたえることを主観的に経験している.このことは,知覚の形成においてはボトムアップの入力だけでなくトップダウンの入力も関連することを示す.しかし,知覚をつかさどる神経回路に対するトップダウンの入力の役割は十分に検証されていない.げっ歯類において第1体性感覚野に対する高次の脳領域はいくつか存在し,そのひとつとして第2運動野がある.第2運動野は大脳皮質の前方に存在し,価値にもとづく意思決定1) や随意運動2) といった高次の脳機能との関連が示唆されてきた.第2運動野と第1体性感覚野とは解剖学的に互いに結合していることから,第1体性感覚野における神経活動は第2運動野からの入力により制御されていると考えられる.そこで,この研究においては,知覚に対するトップダウンの入力の影響を明らかにするため,第2運動野および第1体性感覚野から構成される神経回路の解剖学的および生理学的な同定を試みた.さらに,光遺伝学的な手法を用いた行動実験により,マウスの知覚行動に対する第2運動野から第1体性感覚野へのトップダウンの入力の役割について調べた.
1.第1体性感覚野と第2運動野は機能的に相互に連絡する
マウスの後肢への刺激により第1体性感覚野および第2運動野が活性化されるかどうか検討するため,膜電位の変化により蛍光強度の変化する色素を用いて膜電位イメージング実験を行った.大脳皮質の広範囲な領域から神経活動を記録したところ,後肢への刺激によりまず第1体性感覚野が活動し,それにひきつづき第2運動野が活動することが見い出された.第1体性感覚野と第2運動野とが機能的に相互に連絡しているかどうか検討するため,それぞれの部位における神経活動を薬理学的に抑制した.その結果,第1体性感覚野を抑制したときは第2運動野における神経活動において早い成分が,第2運動野を抑制したときは第1体性感覚野における神経活動において遅い成分が,それぞれ減少した.これらの結果から,まず,後肢からのボトムアップの入力が第1体性感覚野を活性化し,ついで,第1体性感覚野が第2運動野を活性化し,そして,第2運動野がふたたび第1体性感覚野を活性化する,という反響回路の存在が示された(図1a).この反響回路の解剖学的な特徴を調べるため,第1体性感覚野あるいは第2運動野に順行性の神経トレーサーを注入した.その結果,第1体性感覚野から第2運動野へ,および,第2運動野から第1体性感覚野へ,それぞれ投射する神経軸索が確認された.第2運動野から第1体性感覚野へ投射する神経軸索は深層および表層において密であった(図1b).この投射パターンは典型的なトップダウンの入力の投射パターンと同様であった3).
2.第2運動野から第1体性感覚野へのトップダウンの入力は大脳皮質の深層および表層へと伝達される
神経トレーサーを用いた実験から,第2運動野からのトップダウンの入力は第1体性感覚野の深層および表層を活性化すると考えられた.これを検討するため,16点の記録部位が一列にならんだ電極によるマルチユニット記録法を用いて,第1体性感覚野のすべての層(第1層~第6層)から活動電位の応答を記録した.この手法により,1つの記録部位から複数の単一のニューロンの活動電位を記録することができる.後肢への刺激はすべての層において早発性の神経活動および遅発性の神経活動を誘起した.早発性の神経活動は第4層においてもっとも早くピークに達した.これは視床からのボトムアップの入力により誘起された神経活動であると考えられた.一方,遅発性の神経活動は深層から表層という順に生じた.膜電位イメージング実験において,第1体性感覚野における神経活動において遅い成分は第2運動野の神経活動に依存していたことから,この遅発性の神経活動は第2運動野における神経活動により誘起されると考えられた.これを検証するため,第2運動野における神経活動を薬理学的に抑制したところ,遅発性の神経活動だけが抑制された.この結果から,遅発性の神経活動が第2運動野からのトップダウンの入力に起因することが示された.遅発性の神経活動において第5層における神経活動の大きさはどの層よりも顕著であったが,これは第2運動野から第1体性感覚野への神経軸索の投射パターンから予測される神経活動とは一致しなかった.
3.第2運動野から第1体性感覚野へのトップダウンの入力は第5層の錐体細胞の樹状突起におけるCa2+スパイクおよび細胞体における持続的な発火活動を誘起する
第2運動野からの神経軸索は第1体性感覚野の深層および表層に投射していたにもかかわらず,第2運動野における神経活動に由来する遅発性の神経活動は第5層において顕著であった.この理由のひとつとして,第2運動野から第1体性感覚野へのシナプス入力は第5層に集中している可能性が考えられた.そこで,第2運動野から第1体性感覚野への興奮性のシナプス入力の強度を推定するため,第1体性感覚野において記録した細胞外電位から電流源密度解析を行った.その結果,マルチユニット記録法により得られた結果と同じように,後肢への刺激によりすべての層において早発性のシナプス入力および遅発性のシナプス入力が生じた.遅発性のシナプス入力の大きさは第1体性感覚野の深層および表層において顕著であり,これは第2運動野からの投射パターンと一致していた.
第1体性感覚野の深層および表層におけるシナプス入力はおよそ30ミリ秒のあいだ重複していた.第5層の錐体細胞は表層まで樹状突起を伸長していることを考えると,この細胞は30ミリ秒のあいだに深層にある基底の樹状突起および表層にある遠位の樹状突起により第2運動野からの入力をうけ,これを統合している可能性が考えられた.これまでの研究により,第5層の錐体細胞が30ミリ秒のあいだにこれらの入力を統合することにより遠位の樹状突起においてCa2+スパイクの発生することが知られている4).また,このCa2+スパイクにより細胞体においては持続的な発火活動が誘起される5,6).そこで,神経トレーサー実験の結果とマルチユニット記録における結果の不一致は,第1体性感覚野の深層および表層へのシナプス入力が第5層の錐体細胞の樹状突起においてCa2+スパイクを誘起し,これにより持続的な発火活動がひき起こされたためであるとの仮説をたてた.これを検証するため,2光子イメージング法を用いて第5層の錐体細胞の遠位の樹状突起からのCa2+応答を記録したところ,後肢への刺激により“小さくて早いCa2+応答”および“大きくて遅いCa2+応答”が観察された.薬理学的な実験により,この大きくて遅いCa2+応答は第2運動野の神経活動に依存的であることが示され,また,このCa2+応答はCa2+スパイクであることが示唆された.この結果およびマルチユニット記録における薬理学的な実験の結果から,第2運動野からの入力が第5層の錐体細胞の樹状突起においてCa2+スパイクを誘起し,これにより細胞体において持続的な発火活動が生じたことが示された.
4.第2運動野からのトップダウンの入力を抑制すると第1体性感覚野における神経活動が減少し触覚にもとづくマウスの行動が変化する
第2運動野からのトップダウンの入力がマウスの知覚行動にあたえる影響について調べるため,後肢への触覚にもとづく行動課題を行った.第2運動野から第1体性感覚野への神経軸索の投射を光遺伝学的に抑制するため,第2運動野のニューロンに光感受性のイオンポンプを発現させたマウスを用いた.まず,自発性の場所選好試験を行った.この実験では,箱のなかに2種類の異なる床面,ツルツル床およびザラザラ床を配置した.光を照射されないマウスはどちらか一方の床面に対し滞在時間のかたより(つまり,選好性)を示したが,光の照射により第2運動野から第1体性感覚野への入力を抑制したマウスはその滞在時間のかたよりが減少した.この結果は,光の照射により床面に対する知覚が変化したためと考えられた.つぎに,刺激誘発性の肢動作試験を行った.マウスは後肢を軽く刺激されるとある確率で肢を動かす.光の照射により第2運動野から第1体性感覚野への入力を抑制したとき,刺激された肢を動かす確率がどう変化するかを検討した.このとき,第1体性感覚野にて生じる第2運動野からの入力に由来する遅発性の神経活動を選択的に抑制した.その結果,光を照射しない条件と比べ肢を動かす確率は低下し,後肢への刺激により誘発される遅発性の神経活動が知覚の形成に必要であることが示された.
皮膚感覚の知覚が第2運動野から第1体性感覚野へのトップダウンの入力の抑制により変容しているなら,その知覚をもとにした高次機能を示す行動も変化する可能性が考えられた.これを検証するため,Y字型迷路による床面弁別行動試験を行った.この実験では,Y字型迷路の分岐点の手前にツルツル床あるいはザラザラ床を配置し,おのおのの触感に対応した方向に報酬を提示した(たとえば,ザラザラ床の場合には左の方向に報酬が提示される).マウスは訓練により床面の触感と報酬が提示される方向との関係を約80%の正解率まで学習した.これらのマウスを用い,光の照射により第2運動野から第1体性感覚野への入力を抑制すると,正解率は約65%まで低下した.一方,光の照射により,運動,視覚の能力,不安様の行動などに影響はみられなかった.これらの行動実験より,正常な知覚行動には第2運動野から第1体性感覚野へのトップダウンの入力が必須であることが明らかにされた.
おわりに
この研究において,マウスの体性感覚において第2運動野から第1体性感覚野へのトップダウンの入力の神経回路およびその生理的な役割が明らかにされた.具体的には,1)視床からのボトムアップの入力により第1体性感覚野において早発性の神経活動が生じ,これが第2運動野へと伝達される.活性化された第2運動野は第1体性感覚野における遅発性の神経活動を誘起する.2)第2運動野からのトップダウンの入力は第1体性感覚野の深層および表層へと投射され,これらの入力により第5層の錐体細胞の樹状突起においてCa2+スパイクが発生し,細胞体において持続的な発火活動が生じる.3)第2運動野からの入力を第1体性感覚野において光遺伝学的に抑制するとマウスの正確な知覚行動が変化する.
これまでも,第1次感覚野において感覚刺激により誘起される遅発性の神経活動については報告されており,この神経活動の振幅と行動課題の成績とのあいだに強い正の相関のあることが示唆されていた7-11).しかし,この遅発性の神経活動が知覚に必須なのか,それとも,知覚の体験による余波としての神経活動なのかは明らかにされていなかった.最近の報告によると,マウスを用いたヒゲへの刺激の弁別課題において,第1体性感覚野における抑制性のニューロンを光遺伝学的に活性化することにより遅発性の神経活動を抑制すると,マウスはあたかもヒゲへの刺激を知覚していないような行動を示す10).この結果および今回の結果は,遅発性の神経活動は知覚に必須であることを示す.さらに今回,この遅発性の神経活動の実体は第2運動野からのトップダウンの入力による第5層の錐体細胞の樹状突起におけるCa2+スパイクとそれにつづく持続した発火活動であることが明らかにされた.
われわれはどのような神経活動により知覚を体験するのか.単一のニューロンのレベルでの知覚モデルのひとつとして,連合入力モデルがある(図2a).われわれが知覚するためには,視床に由来するボトムアップの入力とは別に,注意や予測といった脳内の活動による“内因性”のトップダウンの入力も必要とするというモデルである.ボトムアップの入力および内因性のトップダウンの入力が脳のある領域において短い時間のあいだ連合することにより樹状突起においてCa2+スパイクが発生し,それにつづく細胞体における持続的な発火活動が知覚をひき起こす.もしこの仮説が正しければ,われわれは“なにかに注意しなければなにも感じない”ことにもなる.しかし実際には,とくに注意せずぼんやりしている状態でも皮膚の感覚を知覚することは可能であり,この仮説ではわれわれの実体験をすべて説明することはできない.今回,見い出された反響回路では,肢からの情報が第1体性感覚野を経由して第2運動野において“外因性”のトップダウンの入力に変換され第1体性感覚野にフィードバックされる(図2b).筆者らは,従来の連合入力モデルと同様に,この外因性のトップダウンの入力のみで樹状突起においてCa2+スパイクをひき起こす機能を見い出した.この反響回路が,われわれがとくに注意などしていない状態においても知覚の形成を可能にする基本的な神経回路として動作している可能性がある.脳はそのときどきの状態により,連合入力モデルによる神経回路と反響回路とを使い分けていると考えられる.
文 献
- Sul, J. H., Jo, S., Lee, D. et al.: Role of rodent secondary motor cortex in value-based action selection. Nat. Neurosci., 14, 1202-1208 (2011)[PubMed]
- Murakami, M., Vicente, M. I., Costa, G. M. et al.: Neural antecedents of self-initiated actions in secondary motor cortex. Nat. Neurosci., 17, 1574-1582 (2014)[PubMed]
- Felleman, D. J. & Van Essen, D. C.: Distributed hierarchical processing in the primate cerebral cortex. Cereb. Cortex, 1, 1-47 (1991)[PubMed]
- Ledergerber, D. & Larkum, M. E.: The time window for generation of dendritic spikes by coincidence of action potentials and EPSPs is layer specific in somatosensory cortex. PLoS One, 7, e33146 (2012)[PubMed]
- Larkum, M. E., Zhu, J. J. & Sakmann, B.: A new cellular mechanism for coupling inputs arriving at different cortical layers. Nature, 398, 338-341 (1999)[PubMed]
- Xu, N. L., Harnett, M. T., Williams, S. R. et al.: Nonlinear dendritic integration of sensory and motor input during an active sensing task. Nature, 492, 247-251 (2012)[PubMed]
- Cauller, L.: Layer I of primary sensory neocortex: where top-down converges upon bottom-up. Behav. Brain Res., 71, 163-170 (1995)[PubMed]
- Del Cul, A., Baillet, S. & Dehaene, S.: Brain dynamics underlying the nonlinear threshold for access to consciousness. PLoS Biol, 5, e260 (2007)[PubMed]
- Gilbert, C. D. & Sigman, M.: Brain states: top-down influences in sensory processing. Neuron, 54, 677-696 (2007)[PubMed]
- Sachidhanandam, S., Sreenivasan, V., Kyriakatos, A. et al.: Membrane potential correlates of sensory perception in mouse barrel cortex. Nat. Neurosci., 16, 1671-1677 (2013)[PubMed]
- Super, H., Spekreijse, H. & Lamme, V. A.: Two distinct modes of sensory processing observed in monkey primary visual cortex (V1). Nat. Neurosci., 4, 304-310 (2001)[PubMed]
著者プロフィール
略歴:2006年東京薬科大学大学院生命科学研究科 修了,同年 米国New York Medical College研究員,2010年 理化学研究所脳科学総合研究センター 研究員を経て,2015年より山梨大学医学部 特任助教.
研究テーマ:生きている動物の脳における単一のニューロンの特性.
抱負:脳切片標本から得られた知見が生きた脳においても適応できるのかといった問題や,単一のニューロンの活動と動物の行動や学習といった脳の高次機能との因果関係について明らかにしたい.
鈴木 崇之(Takayuki Suzuki)
理化学研究所脳科学総合研究センター 研究員.
村山 正宜(Masanori Murayama)
理化学研究所脳科学総合研究センター チームリーダー.
研究室URL:http://murayama-lab.brain.riken.jp/index_jp.html
© 2015 真仁田 聡・鈴木崇之・村山正宜 Licensed under CC 表示 2.1 日本
