高等植物の光化学系I–光捕集アンテナI超複合体におけるエネルギー伝達経路の構造基盤
菅 倫寛・沈 建仁
(岡山大学大学院自然科学研究科 地球生命物質科学専攻構造生物学分野)
email:沈 建仁
DOI: 10.7875/first.author.2015.074
Structural basis for energy transfer pathways in the plant PSI-LHCI supercomplex.
Xiaochun Qin, Michihiro Suga, Tingyun Kuang, Jian-Ren Shen
Science, 348, 989-995 (2015)
光化学系I複合体は光エネルギーを利用して水からの電子を用いて糖の合成に必要な還元力となるNADPHをつくりだす.高等植物の光化学系Iコアには光捕集アンテナIが会合して光化学系I-光捕集アンテナI超複合体を形成し,100%に近いエネルギーの伝達および変換の効率を実現している.光合成における高い効率での太陽光の利用の基本原理を理解することはエネルギー問題や環境問題をかかえる現代社会においてきわめて重要な課題である.今回,筆者らは,高等植物に由来する600 kDにもおよぶ光化学系I-光捕集アンテナI超複合体のX線結晶構造解析を行い,その構造を分解能2.8Åで決定した.これにより,高等植物の光化学系I-光捕集アンテナI超複合体の詳細な構造,とくに,光捕集アンテナIを構成する4つのLhcaサブユニットにおいて秩序正しく配置されたクロロフィル,カロテノイド,脂質分子,水分子などの構造基盤を明らかにし,光エネルギーの捕集および伝達の経路,および,光阻害に対する防御機構を提唱した.
植物や各種の藻類による酸素発生型の光合成は太陽光を利用してCO2と水から有機物を合成する反応であり,その副産物であるO2はわれわれ人類を含む地球上すべての好気性生物の生存をささえている.光合成においては太陽光を駆動力として,光化学系II複合体は水を酸素,プロトン,電子に分解し,光化学系I複合体はこの電子を利用してNADP+を還元し糖の合成に必要な還元力となるNADPHをつくりだす.地球上ではじめて酸素発生型の光合成能を獲得したシアノバクテリアはおよそ30億年まえに出現し,以来その進化の過程において光合成の効率を向上させてきた.光化学系II複合体および光化学系I複合体における最大の進化はアンテナ系タンパク質複合体の獲得であり,シアノバクテリアの光化学系I複合体は三量体として存在するのに対し1),高等植物の光化学系I複合体は単量体として存在し光捕集アンテナIと600 kDにもおよぶ超複合体を形成する2).光化学系I-光捕集アンテナI超複合体における光エネルギーの伝達および変換の効率はほぼ100%とされており3),その高い効率での光エネルギーの利用機構の解明は太陽光発電の効率の向上や人工光合成の研究への応用などにもつながると期待されている.
これまでの分光学的および生化学的な研究から,植物の光捕集アンテナIは4つの膜タンパク質Lhca1~Lhca4から構成される複合体であり,クロロフィルa,クロロフィルb,複数種のカロテノイドを適切に配置してエネルギーを高い効率で伝達するだけでなく,過度に蓄積されたエネルギーを熱として散逸させるクエンチングのしくみももつことが明らかにされている.しかし,光化学系I-光捕集アンテナI超複合体は巨大な膜タンパク質複合体であるため,これまでに解析された高等植物の光化学系I-光捕集アンテナI超複合体の結晶構造の分解能は3.3Åにとどまり2,4,5),光捕集アンテナIの発揮するこれらの機能を理解するには不十分であったため,原子分解能での結晶構造が待ち望まれていた.
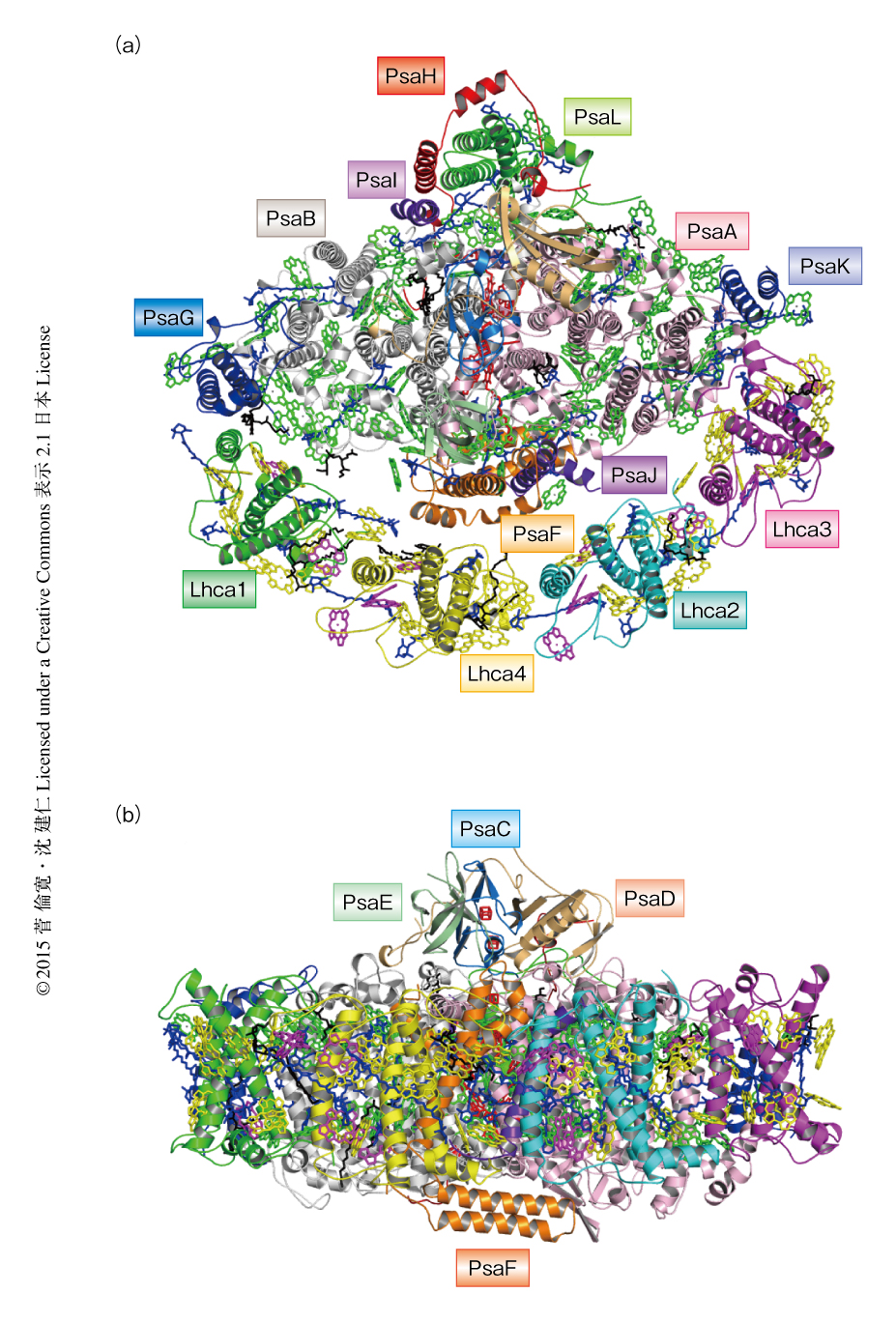
エンドウマメの葉から単離した光化学系I-光捕集アンテナI超複合体を結晶化し,大型放射光施設SPring-8の高輝度X線を用いて結晶構造を分解能2.8Åで決定した(PDB ID:4XK8).その結果,光化学系I-光捕集アンテナI超複合体はLhca1~Lhca4の4つのサブユニットを含む計16つのサブユニットから構成され,長さ140Åの親骨をもつ扇子のような構造をとることがわかった(図1).光化学系Iコア複合体のうち,PsaHはちょうど扇子の要となり扇子の中骨に位置する光化学系Iコアを固定しており,Lhca1~Lhca4は2つのヘテロ二量体Lhca1-Lhca4およびLhca2-Lhca3を形成し,それらのあいだでさらにヘテロ二量体構造をとりPsaF,PsaJ,PsaK,PsaGの位置する側に扇子の扇面のように結合していた.高分解能での構造解析により,光化学系I-光捕集アンテナI超複合体に含まれる,143個のクロロフィルa,12個のクロロフィルb,35個のカロテノイド(26個のβカロテン,5個のルテイン,4個のビオラキサンチン),10個の脂質分子(6個のホスファチジルグリセロール,3個のモノガラクトシルジアシルグリセロール,1個のジガラクトシルジアシルグリセロール),3個のFe4S4クラスター,2個のフィロキノンの位置および配置が正確に決定された.とりわけ,クロロフィルおよびカロテノイドの数はこれまでの構造解析による報告とは大きく異なっていたが2,4,5),今回の結果は,結晶化された標品を用いたHPLC分析の結果やこれまでの生化学的な実験の報告ともよく一致したことから,光化学系I-光捕集アンテナI超複合体のほぼすべての集光性色素を同定することに成功したと考えられた.
光捕集アンテナIには,45個のクロロフィルa,12個のクロロフィルb,4個のβカロテン,5個のルテイン,4個のビオラキサンチンが結合していた.光捕集アンテナIにおいてクロロフィルはチラコイド膜に平行した2つの層に分布しており,ひとつの層はストロマの近くに,もうひとつの層はルーメンの近くに位置していた.ストロマ側の層には29個のクロロフィルaおよび7個のクロロフィルbが分布しており,となりあうクロロフィルのMg2+とMg2+とのあいだの平均距離は11.3Åであった.ルーメン側の層はストロマ側の層よりもクロロフィルの密度が低く,16個のクロロフィルaと5個のクロロフィルbを含む2つのクラスターを形成しており,Mg2+とMg2+とのあいだの平均距離はクラスターの内部では10.6Å,クラスターのあいだでは19.7Åであった.
高分解能の結晶構造が既知である光捕集アンテナIIと比較したところ6),向きの異なるクロロフィル,あるいは,まったく別の部位に結合しているクロロフィルがいくつかみつかったが,それらのほとんどは光化学系Iコアと光捕集アンテナIとの空隙の部分か,隣接するLhca1~Lhca4サブユニットのあいだのいずれかに位置していた.光捕集アンテナIに特有のクロロフィルは,光化学系I-光捕集アンテナI超複合体における高い効率での光エネルギーの利用機構と密接に関係していると考えられた.
光捕集アンテナIのLhca1~Lhca4サブユニットはそれぞれが3~4個のカロテノイドと結合していた.高分解能の結晶構造が既知である光捕集アンテナIIでは4つのカロテノイドの結合部位がみつかっており6),膜貫通ヘリックスAおよび膜貫通ヘリックスBの形成するくぼみにルテインと結合するL1部位およびL2部位,膜貫通ヘリックスCの近傍にネオキサンチンと結合するN1部位,隣接する光捕集アンテナIIとの境界にビオラキサンチンと結合するV1部位が存在する.光捕集アンテナIにおいては,いずれのLhcaサブユニットにおいてもL1部位にルテイン,L2部位にビオラキサンチン,N1部位にβカロテンが結合していたが,V1部位にはカロテノイドは何もみつからず,Lhca1のみ新規のL4部位にルテインが結合していた.
Lhca1~Lhca4サブユニットに共通してみつかった3つのカロテノイドの結合部位のうち,L1部位のみが光捕集アンテナIIおよびCP29 7) と同じくルテインと結合していた.光捕集アンテナIIにおける研究により,L1部位に結合するルテインは非光化学的なクエンチングの機能をはたすことがわかっていることから8),L1部位およびその周辺の構造を比較したところ,光捕集アンテナIのL1部位にみつかったルテインと周囲のクロロフィルaのクラスターは光捕集アンテナIIおよびCP29と向きおよび位置関係が非常によく似ていることがわかった6,7).このことから,光捕集アンテナIにおいてもL1部位に結合したルテインは近傍のクロロフィルaのクラスターと共役して過剰に蓄積したエネルギーを散逸させる役割をはたすことが示唆された.
L2部位について,光捕集アンテナIIにおいてはルテイン,CP29および光捕集アンテナIにおいてはビオラキサンチンが結合していた.古くからキサントフィル回路として知られている光阻害に対する防御機構においては,光捕集アンテナIIのビオラキサンチンが脱エポキシ化反応によりゼアキサンチンへと変化して励起されたクロロフィルを消光させると考えられている9).これまでの研究から,光捕集アンテナIも光阻害に対する同様の防御機構を備えている可能性は指摘されていたが,つい最近の,光捕集アンテナIにおけるゼアキサンチンによる光エネルギーの捕集の制御に関する報告まで10),多くの研究者は光捕集アンテナIにおいてキサントフィル回路は機能していないと考えていた.今回の光化学系I-光捕集アンテナI超複合体の結晶構造は,光捕集アンテナIにおいてLhca1~Lhca4サブユニットのL2部位はそれぞれ異なる機能を発揮する直接的な証拠を示した.Lhca1~Lhca4サブユニットのそれぞれにおいてL2部位をとりかこむクロロフィルは異なる水素結合ネットワークをもち,Lhca2およびLhca4においては光捕集アンテナIIおよびCP29の水素結合ネットワークとよく似ていた一方6,7),Lhca1およびLhca3においては似ていなかった.これらの水素結合ネットワークの違いは,光エネルギーの捕集およびクエンチングにおける役割の違いを反映している可能性が考えられた.
今回の光化学系I-光捕集アンテナI超複合体の結晶構造から明らかにされた光化学系Iコアと光捕集アンテナIとのあいだに存在する集光性色素の位置関係にもとづき,これまでの分光学的な研究による知見をふまえ,光捕集アンテナIの捕集した光エネルギーを光化学系Iコアへと伝達するエネルギー伝達経路の候補を提唱した.光捕集アンテナIの特徴として,赤色型とよばれるエネルギー準位の低いクロロフィル二量体が存在し,それらがエネルギーの伝達に重要な役割をはたしていることがわかっていたが,結晶構造から,光捕集アンテナIの捕集したほぼすべての光エネルギーは赤色型であるクロロフィル二量体を経由し高い効率で光化学系Iコアへと伝達されることがわかった.結晶構造から,光捕集アンテナIから光化学系Iコアへのもっとも可能性の高いエネルギー伝達経路として,1Bs経路,1Fl経路,2Js経路,3As/3Al経路が同定された.
1Bs経路はLhca1に集められたエネルギーをPsaBに伝達するストロマの側の経路である.1Fl経路はLhca1からPsaFのルーメンの側につながる経路である.Lhca4と光化学系Iコアとは距離が離れており,Lhca4に集められたエネルギーを光化学系Iコアに直接に伝達するのはむずかしいようであるが,この1Fl経路を利用してこのギャップを克服しているように思われた.2Js経路はLhca2とPsaJとを結ぶ経路である.Lhca2はエネルギーをPsaJのストロマの側に直接に伝達するには距離が離れているため,この2Js経路はそれほど機能していないのかもしれない.Lhca3はPsaAのストロマの側と3As経路を形成する.この3As経路はLhca3からのエネルギー伝達経路としてとくに効率よく機能していると考えられた.Lhca3のルーメンの側もPsaAと近接しており,3Al経路を形成する.さらに,Lhca3はPsaKのルーメンの側にエネルギーを伝達することが可能な距離にあり,3Kl経路を形成する.
この研究がScience誌に出版された当日,競争相手からメールが届いた.“おめでとう.じつはわれわれも,ほぼ同じ程度の分解能の結晶構造を得ることに成功していて,投稿の準備を進めているところだ”.以前から競争関係にあることは想定していたが,そこまで進展しているとはまったく予想していなかった.本当に気の毒に思うが,一歩違えばまったく逆の立場だった可能性もあるわけで,今回の研究から,とくに膜タンパク質を対象とした国際競争の厳しさを肌で感じた.そのような事情もあり,この研究においては600 kDの超複合体の構造をわずか2週間で組み上げた.構造解析にはベストをつくしたが,いくつかの弱い電子密度を帰属できないままにしてしまったことは心残りである.厳しい競争のなかアウトプットがもとめられがちになる傾向があるが,スピーディーさを維持しつつも妥協せずていねいな研究を心がけたいと思う.
略歴:2009年 大阪大学大学院理学研究科 修了,大阪大学蛋白質研究所 研究員,米国Oregon Health & Science大学 研究員を経て,岡山大学大学院自然科学研究科 特任助教(現 助教).
研究テーマ:膜タンパク質の構造生物学.
沈 建仁(Jian-Ren Shen)
岡山大学大学院自然科学研究科 教授.
© 2015 菅 倫寛・沈 建仁 Licensed under CC 表示 2.1 日本
(岡山大学大学院自然科学研究科 地球生命物質科学専攻構造生物学分野)
email:沈 建仁
DOI: 10.7875/first.author.2015.074
Structural basis for energy transfer pathways in the plant PSI-LHCI supercomplex.
Xiaochun Qin, Michihiro Suga, Tingyun Kuang, Jian-Ren Shen
Science, 348, 989-995 (2015)
要 約
光化学系I複合体は光エネルギーを利用して水からの電子を用いて糖の合成に必要な還元力となるNADPHをつくりだす.高等植物の光化学系Iコアには光捕集アンテナIが会合して光化学系I-光捕集アンテナI超複合体を形成し,100%に近いエネルギーの伝達および変換の効率を実現している.光合成における高い効率での太陽光の利用の基本原理を理解することはエネルギー問題や環境問題をかかえる現代社会においてきわめて重要な課題である.今回,筆者らは,高等植物に由来する600 kDにもおよぶ光化学系I-光捕集アンテナI超複合体のX線結晶構造解析を行い,その構造を分解能2.8Åで決定した.これにより,高等植物の光化学系I-光捕集アンテナI超複合体の詳細な構造,とくに,光捕集アンテナIを構成する4つのLhcaサブユニットにおいて秩序正しく配置されたクロロフィル,カロテノイド,脂質分子,水分子などの構造基盤を明らかにし,光エネルギーの捕集および伝達の経路,および,光阻害に対する防御機構を提唱した.
はじめに
植物や各種の藻類による酸素発生型の光合成は太陽光を利用してCO2と水から有機物を合成する反応であり,その副産物であるO2はわれわれ人類を含む地球上すべての好気性生物の生存をささえている.光合成においては太陽光を駆動力として,光化学系II複合体は水を酸素,プロトン,電子に分解し,光化学系I複合体はこの電子を利用してNADP+を還元し糖の合成に必要な還元力となるNADPHをつくりだす.地球上ではじめて酸素発生型の光合成能を獲得したシアノバクテリアはおよそ30億年まえに出現し,以来その進化の過程において光合成の効率を向上させてきた.光化学系II複合体および光化学系I複合体における最大の進化はアンテナ系タンパク質複合体の獲得であり,シアノバクテリアの光化学系I複合体は三量体として存在するのに対し1),高等植物の光化学系I複合体は単量体として存在し光捕集アンテナIと600 kDにもおよぶ超複合体を形成する2).光化学系I-光捕集アンテナI超複合体における光エネルギーの伝達および変換の効率はほぼ100%とされており3),その高い効率での光エネルギーの利用機構の解明は太陽光発電の効率の向上や人工光合成の研究への応用などにもつながると期待されている.
これまでの分光学的および生化学的な研究から,植物の光捕集アンテナIは4つの膜タンパク質Lhca1~Lhca4から構成される複合体であり,クロロフィルa,クロロフィルb,複数種のカロテノイドを適切に配置してエネルギーを高い効率で伝達するだけでなく,過度に蓄積されたエネルギーを熱として散逸させるクエンチングのしくみももつことが明らかにされている.しかし,光化学系I-光捕集アンテナI超複合体は巨大な膜タンパク質複合体であるため,これまでに解析された高等植物の光化学系I-光捕集アンテナI超複合体の結晶構造の分解能は3.3Åにとどまり2,4,5),光捕集アンテナIの発揮するこれらの機能を理解するには不十分であったため,原子分解能での結晶構造が待ち望まれていた.
1.光化学系I-光捕集アンテナI超複合体の全体構造
エンドウマメの葉から単離した光化学系I-光捕集アンテナI超複合体を結晶化し,大型放射光施設SPring-8の高輝度X線を用いて結晶構造を分解能2.8Åで決定した(PDB ID:4XK8).その結果,光化学系I-光捕集アンテナI超複合体はLhca1~Lhca4の4つのサブユニットを含む計16つのサブユニットから構成され,長さ140Åの親骨をもつ扇子のような構造をとることがわかった(図1).光化学系Iコア複合体のうち,PsaHはちょうど扇子の要となり扇子の中骨に位置する光化学系Iコアを固定しており,Lhca1~Lhca4は2つのヘテロ二量体Lhca1-Lhca4およびLhca2-Lhca3を形成し,それらのあいだでさらにヘテロ二量体構造をとりPsaF,PsaJ,PsaK,PsaGの位置する側に扇子の扇面のように結合していた.高分解能での構造解析により,光化学系I-光捕集アンテナI超複合体に含まれる,143個のクロロフィルa,12個のクロロフィルb,35個のカロテノイド(26個のβカロテン,5個のルテイン,4個のビオラキサンチン),10個の脂質分子(6個のホスファチジルグリセロール,3個のモノガラクトシルジアシルグリセロール,1個のジガラクトシルジアシルグリセロール),3個のFe4S4クラスター,2個のフィロキノンの位置および配置が正確に決定された.とりわけ,クロロフィルおよびカロテノイドの数はこれまでの構造解析による報告とは大きく異なっていたが2,4,5),今回の結果は,結晶化された標品を用いたHPLC分析の結果やこれまでの生化学的な実験の報告ともよく一致したことから,光化学系I-光捕集アンテナI超複合体のほぼすべての集光性色素を同定することに成功したと考えられた.
2.光捕集アンテナIにおけるクロロフィルの配置とその機能
光捕集アンテナIには,45個のクロロフィルa,12個のクロロフィルb,4個のβカロテン,5個のルテイン,4個のビオラキサンチンが結合していた.光捕集アンテナIにおいてクロロフィルはチラコイド膜に平行した2つの層に分布しており,ひとつの層はストロマの近くに,もうひとつの層はルーメンの近くに位置していた.ストロマ側の層には29個のクロロフィルaおよび7個のクロロフィルbが分布しており,となりあうクロロフィルのMg2+とMg2+とのあいだの平均距離は11.3Åであった.ルーメン側の層はストロマ側の層よりもクロロフィルの密度が低く,16個のクロロフィルaと5個のクロロフィルbを含む2つのクラスターを形成しており,Mg2+とMg2+とのあいだの平均距離はクラスターの内部では10.6Å,クラスターのあいだでは19.7Åであった.
高分解能の結晶構造が既知である光捕集アンテナIIと比較したところ6),向きの異なるクロロフィル,あるいは,まったく別の部位に結合しているクロロフィルがいくつかみつかったが,それらのほとんどは光化学系Iコアと光捕集アンテナIとの空隙の部分か,隣接するLhca1~Lhca4サブユニットのあいだのいずれかに位置していた.光捕集アンテナIに特有のクロロフィルは,光化学系I-光捕集アンテナI超複合体における高い効率での光エネルギーの利用機構と密接に関係していると考えられた.
3.光捕集アンテナIにおけるカロテノイドの配置とその機能
光捕集アンテナIのLhca1~Lhca4サブユニットはそれぞれが3~4個のカロテノイドと結合していた.高分解能の結晶構造が既知である光捕集アンテナIIでは4つのカロテノイドの結合部位がみつかっており6),膜貫通ヘリックスAおよび膜貫通ヘリックスBの形成するくぼみにルテインと結合するL1部位およびL2部位,膜貫通ヘリックスCの近傍にネオキサンチンと結合するN1部位,隣接する光捕集アンテナIIとの境界にビオラキサンチンと結合するV1部位が存在する.光捕集アンテナIにおいては,いずれのLhcaサブユニットにおいてもL1部位にルテイン,L2部位にビオラキサンチン,N1部位にβカロテンが結合していたが,V1部位にはカロテノイドは何もみつからず,Lhca1のみ新規のL4部位にルテインが結合していた.
Lhca1~Lhca4サブユニットに共通してみつかった3つのカロテノイドの結合部位のうち,L1部位のみが光捕集アンテナIIおよびCP29 7) と同じくルテインと結合していた.光捕集アンテナIIにおける研究により,L1部位に結合するルテインは非光化学的なクエンチングの機能をはたすことがわかっていることから8),L1部位およびその周辺の構造を比較したところ,光捕集アンテナIのL1部位にみつかったルテインと周囲のクロロフィルaのクラスターは光捕集アンテナIIおよびCP29と向きおよび位置関係が非常によく似ていることがわかった6,7).このことから,光捕集アンテナIにおいてもL1部位に結合したルテインは近傍のクロロフィルaのクラスターと共役して過剰に蓄積したエネルギーを散逸させる役割をはたすことが示唆された.
L2部位について,光捕集アンテナIIにおいてはルテイン,CP29および光捕集アンテナIにおいてはビオラキサンチンが結合していた.古くからキサントフィル回路として知られている光阻害に対する防御機構においては,光捕集アンテナIIのビオラキサンチンが脱エポキシ化反応によりゼアキサンチンへと変化して励起されたクロロフィルを消光させると考えられている9).これまでの研究から,光捕集アンテナIも光阻害に対する同様の防御機構を備えている可能性は指摘されていたが,つい最近の,光捕集アンテナIにおけるゼアキサンチンによる光エネルギーの捕集の制御に関する報告まで10),多くの研究者は光捕集アンテナIにおいてキサントフィル回路は機能していないと考えていた.今回の光化学系I-光捕集アンテナI超複合体の結晶構造は,光捕集アンテナIにおいてLhca1~Lhca4サブユニットのL2部位はそれぞれ異なる機能を発揮する直接的な証拠を示した.Lhca1~Lhca4サブユニットのそれぞれにおいてL2部位をとりかこむクロロフィルは異なる水素結合ネットワークをもち,Lhca2およびLhca4においては光捕集アンテナIIおよびCP29の水素結合ネットワークとよく似ていた一方6,7),Lhca1およびLhca3においては似ていなかった.これらの水素結合ネットワークの違いは,光エネルギーの捕集およびクエンチングにおける役割の違いを反映している可能性が考えられた.
4.光捕集アンテナIから光化学系Iコアへのエネルギー伝達経路
今回の光化学系I-光捕集アンテナI超複合体の結晶構造から明らかにされた光化学系Iコアと光捕集アンテナIとのあいだに存在する集光性色素の位置関係にもとづき,これまでの分光学的な研究による知見をふまえ,光捕集アンテナIの捕集した光エネルギーを光化学系Iコアへと伝達するエネルギー伝達経路の候補を提唱した.光捕集アンテナIの特徴として,赤色型とよばれるエネルギー準位の低いクロロフィル二量体が存在し,それらがエネルギーの伝達に重要な役割をはたしていることがわかっていたが,結晶構造から,光捕集アンテナIの捕集したほぼすべての光エネルギーは赤色型であるクロロフィル二量体を経由し高い効率で光化学系Iコアへと伝達されることがわかった.結晶構造から,光捕集アンテナIから光化学系Iコアへのもっとも可能性の高いエネルギー伝達経路として,1Bs経路,1Fl経路,2Js経路,3As/3Al経路が同定された.
1Bs経路はLhca1に集められたエネルギーをPsaBに伝達するストロマの側の経路である.1Fl経路はLhca1からPsaFのルーメンの側につながる経路である.Lhca4と光化学系Iコアとは距離が離れており,Lhca4に集められたエネルギーを光化学系Iコアに直接に伝達するのはむずかしいようであるが,この1Fl経路を利用してこのギャップを克服しているように思われた.2Js経路はLhca2とPsaJとを結ぶ経路である.Lhca2はエネルギーをPsaJのストロマの側に直接に伝達するには距離が離れているため,この2Js経路はそれほど機能していないのかもしれない.Lhca3はPsaAのストロマの側と3As経路を形成する.この3As経路はLhca3からのエネルギー伝達経路としてとくに効率よく機能していると考えられた.Lhca3のルーメンの側もPsaAと近接しており,3Al経路を形成する.さらに,Lhca3はPsaKのルーメンの側にエネルギーを伝達することが可能な距離にあり,3Kl経路を形成する.
おわりに
この研究がScience誌に出版された当日,競争相手からメールが届いた.“おめでとう.じつはわれわれも,ほぼ同じ程度の分解能の結晶構造を得ることに成功していて,投稿の準備を進めているところだ”.以前から競争関係にあることは想定していたが,そこまで進展しているとはまったく予想していなかった.本当に気の毒に思うが,一歩違えばまったく逆の立場だった可能性もあるわけで,今回の研究から,とくに膜タンパク質を対象とした国際競争の厳しさを肌で感じた.そのような事情もあり,この研究においては600 kDの超複合体の構造をわずか2週間で組み上げた.構造解析にはベストをつくしたが,いくつかの弱い電子密度を帰属できないままにしてしまったことは心残りである.厳しい競争のなかアウトプットがもとめられがちになる傾向があるが,スピーディーさを維持しつつも妥協せずていねいな研究を心がけたいと思う.
文 献
- Jordan, P., Fromme, P., Witt, H. T. et al.: Three-dimensional structure of cyanobacterial photosystem I at 2.5Å resolution. Nature, 411, 909-917 (2001)[PubMed]
- Ben-Shem, A., Frolow, F. & Nelson, N.: Crystal structure of plant photosystem I. Nature, 426, 630-635 (2003)[PubMed]
- Nelson, N.: Plant photosystem I: the most efficient nano-photochmical machine. J. Nanosci. Nanotechonol., 9, 1709-1713 (2009)[PubMed]
- Amunts, A., Drory, O. & Nelson, N.: The structure of a plant photosystem I supercomplex at 3.4Å resolution. Nature, 447, 58-63 (2007)[PubMed]
- Amunts, A., Toporik, H., Borovikova, A. et al.: Structure determination and improved model of plant photosystem I. J. Biol. Chem., 285, 3478-3486 (2010)[PubMed]
- Liu, Z., Yan, H., Wang, K. et al.: Crystal structure of spinach major light-harvesting complex at 2.72Å resolution. Nature, 428, 287-292 (2004)[PubMed]
- Pan, X., Li, M., Wan, T. et al.: Structural insights into energy regulation of light-harvesting complex CP29 from spinach. Nat. Struct. Mol. Biol., 18, 309-315 (2011)[PubMed]
- Ruban, A., Berera, R., Ilioaia, C. et al.: Identification of a mechanism of photoprotective energy dissipation in higher plants. Nature, 450, 575-578 (2007)[PubMed]
- Niyogi, K. K., Grossman, A. R. & Bjorkman, O.: Arabidopsis mutants define a central role for the xanthophyll cycle in the regulation of photosynthetic energy conversion. Plant Cell, 10, 1121-1134 (1998)[PubMed]
- Ballottari, M., Alcocer, M. J. P., D'Andrea, C. et al.: Regulation of photosystem I light harvesting by zeaxanthin. Proc. Natl. Acad. Sci. USA, 111, E2431-E2438 (2014)[PubMed]
著者プロフィール
略歴:2009年 大阪大学大学院理学研究科 修了,大阪大学蛋白質研究所 研究員,米国Oregon Health & Science大学 研究員を経て,岡山大学大学院自然科学研究科 特任助教(現 助教).
研究テーマ:膜タンパク質の構造生物学.
沈 建仁(Jian-Ren Shen)
岡山大学大学院自然科学研究科 教授.
© 2015 菅 倫寛・沈 建仁 Licensed under CC 表示 2.1 日本
